

Application of neural computing methods for interpreting phospholipid fatty acid profiles of natural microbial communities
- PMID: 10653738
- PMCID: PMC91883
- DOI: 10.1128/AEM.66.2.694-699.2000
Application of neural computing methods for interpreting phospholipid fatty acid profiles of natural microbial communities
Abstract
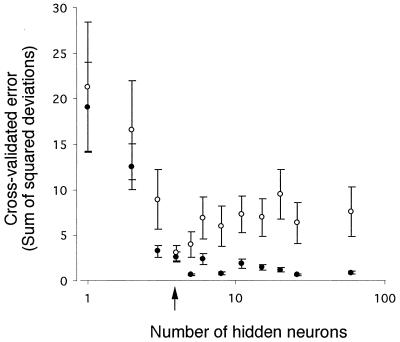
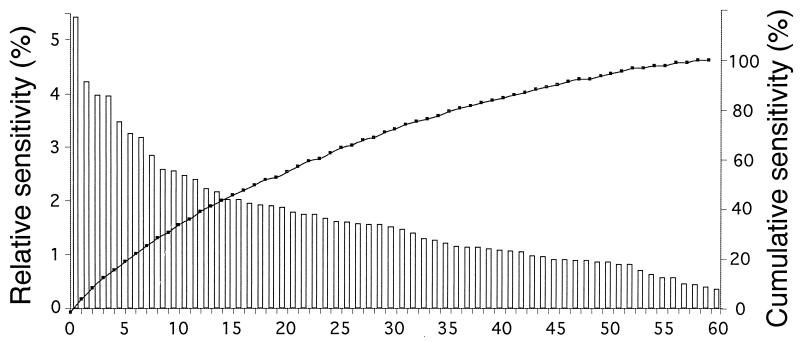
The microbial community compositions of surface and subsurface marine sediments and sediments lining burrows of marine polychaetes and hemichordates from the North Inlet estuary (near Georgetown, S.C. ) were analyzed by comparing ester-linked phospholipid fatty acid (PLFA) profiles with a back-propagating neural network (NN). The NNs were trained to relate PLFA inputs to sediment type outputs (e.g., surface, subsurface, and burrow lining) and worm species (e.g., Notomastus lobatus, Balanoglossus aurantiacus, and Branchyoasychus americana). Sensitivity analysis was used to determine which of the 60 PLFAs significantly contributed to training the NN. The NN architecture was optimized by changing the number of hidden neurons and calculating the cross-validation error between predicted and actual outputs of training and test data. The optimal NN architecture was found to be four hidden neurons with 60-input neurons representing the 60 PLFAs, and four output neurons coding for both sediment types and worm species. Comparison of cross-validation results using NNs and linear discriminant analysis (LDA) revealed that NNs had significantly fewer incorrect classifications (2.7%) than LDA (8.4%). For the NN cross-validation, both sediment type and worm species had 3 incorrect classifications out of 112. For the LDA cross-validation, sediment type and worm species had 7 and 12 incorrect classifications out of 112, respectively. Sensitivity analysis of the trained NNs revealed that 17 fatty acids explained 50% of variability in the data set. These PLFAs were highly different among sediments and burrow types, indicating significant differences in the microbiota.
Figures
References
-
- Acinas S G, Rodriguez-Valera F, Pedros-Alio C. Spatial and temporal variation in marine bacterioplankton diversity as shown by RFLP fingerprinting of PCR amplified 16s rDNA. FEMS Microbiol Ecol. 1997;24:27–40.
-
- Aleksander I, Morton H. An introduction to neural computing. London, England: Chapman & Hall, Ltd.; 1991. Principles and promises; pp. 1–20.
-
- Almeida J S, Sonesson A, Ringelberg D B, White D C. Application of artificial neural networks (ANN) to the detection of Mycobacterium tuberculosis, its antibiotic resistance and prediction of pathogenicity amongst Mycobacterium spp. based on signature lipid biomarkers. Bin Comput Microbiol. 1995;7:53–59.
-
- Baird B H, White D C. Biomass and community structure of the abyssal microbiota determined from the ester-linked phospholipids recovered from Venezuela Basin and Puerto Rico Trench sediment. Mar Geol. 1985;68:217–231. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
