

alpha-Melanocyte-stimulating hormone is contained in nerve terminals innervating thyrotropin-releasing hormone-synthesizing neurons in the hypothalamic paraventricular nucleus and prevents fasting-induced suppression of prothyrotropin-releasing hormone gene expression
- PMID: 10662844
- PMCID: PMC6772359
- DOI: 10.1523/JNEUROSCI.20-04-01550.2000
alpha-Melanocyte-stimulating hormone is contained in nerve terminals innervating thyrotropin-releasing hormone-synthesizing neurons in the hypothalamic paraventricular nucleus and prevents fasting-induced suppression of prothyrotropin-releasing hormone gene expression
Abstract
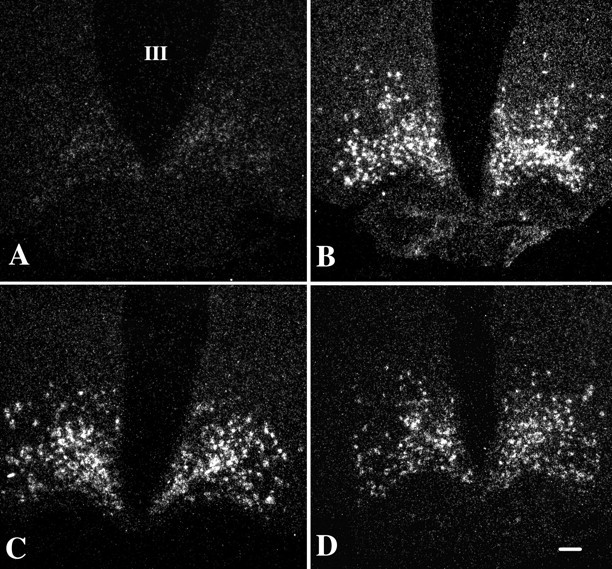
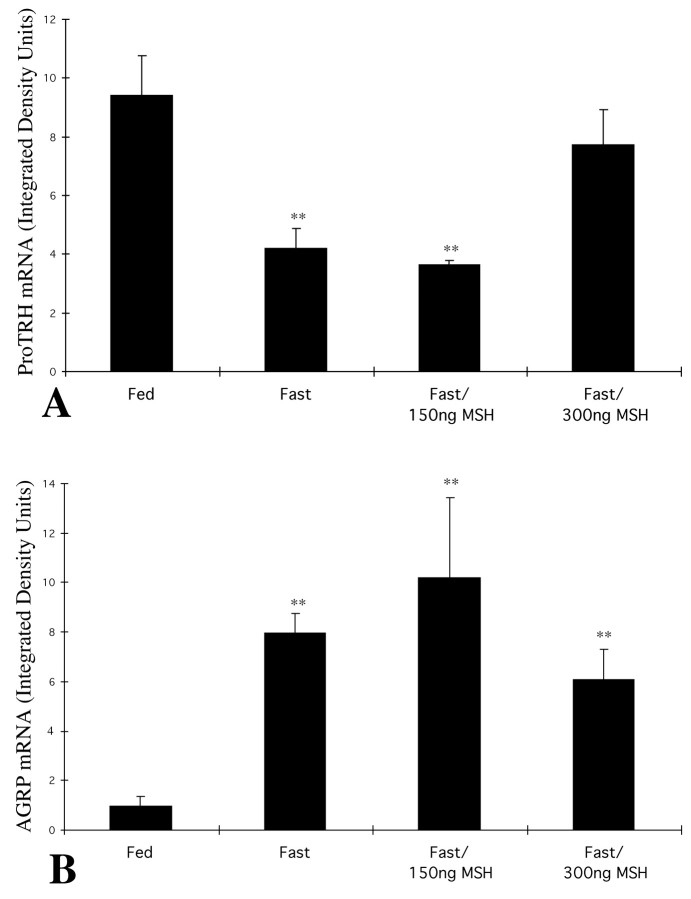
The hypothalamic arcuate nucleus has an essential role in mediating the homeostatic responses of the thyroid axis to fasting by altering the sensitivity of prothyrotropin-releasing hormone (pro-TRH) gene expression in the paraventricular nucleus (PVN) to feedback regulation by thyroid hormone. Because agouti-related protein (AGRP), a leptin-regulated, arcuate nucleus-derived peptide with alpha-MSH antagonist activity, is contained in axon terminals that terminate on TRH neurons in the PVN, we raised the possibility that alpha-MSH may also participate in the mechanism by which leptin influences pro-TRH gene expression. By double-labeling immunocytochemistry, alpha-MSH-IR axon varicosities were juxtaposed to approximately 70% of pro-TRH neurons in the anterior and periventricular parvocellular subdivisions of the PVN and to 34% of pro-TRH neurons in the medial parvocellular subdivision, establishing synaptic contacts both on the cell soma and dendrites. All pro-TRH neurons receiving contacts by alpha-MSH-containing fibers also were innervated by axons containing AGRP. The intracerebroventricular infusion of 300 ng of alpha-MSH every 6 hr for 3 d prevented fasting-induced suppression of pro-TRH in the PVN but had no effect on AGRP mRNA in the arcuate nucleus. alpha-MSH also increased circulating levels of free thyroxine (T4) 2.5-fold over the levels in fasted controls, but free T4 did not reach the levels in fed controls. These data suggest that alpha-MSH has an important role in the activation of pro-TRH gene expression in hypophysiotropic neurons via either a mono- and/or multisynaptic pathway to the PVN, but factors in addition to alpha-MSH also contribute to the mechanism by which leptin administration restores thyroid hormone levels to normal in fasted animals.
Figures



References
-
- Adan RA, Cone RD, Burbach JP, Gispen WH. Differential effects of melanocortin peptides on neural melanocortin receptors. Mol Pharmacol. 1994;46:1182–1190. - PubMed
-
- Beck B, Richy S. Hypothalamic hypocretin/orexin and neuropeptide Y: divergent interaction with energy depletion and leptin. Biochem Biophys Res Commun. 1999;258:119–122. - PubMed
-
- Blake NG, Eckland DJ, Foster OJ, Lightman SL. Inhibition of hypothalamic thyrotropin-releasing hormone messenger ribonucleic acid during food deprivation. Endocrinology. 1991;129:2714–2718. - PubMed
-
- Branchereau P, Van Bockstaele EJ, Chan J, Pickel VM. Ultrastructural characterization of neurons recorded intracellularly in vivo and injected with lucifer yellow: advantages of immunogold-silver vs. immunoperoxidase labeling. Microsc Res Tech. 1995;30:427–436. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources