

Cellular receptor traffic is essential for productive duck hepatitis B virus infection
- PMID: 10666250
- PMCID: PMC111701
- DOI: 10.1128/jvi.74.5.2203-2209.2000
Cellular receptor traffic is essential for productive duck hepatitis B virus infection
Abstract
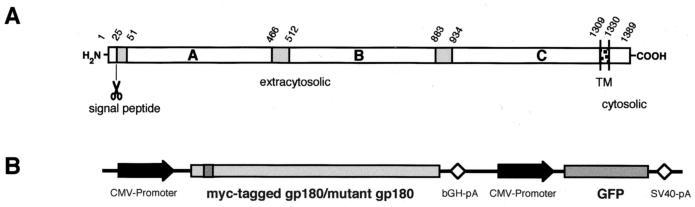
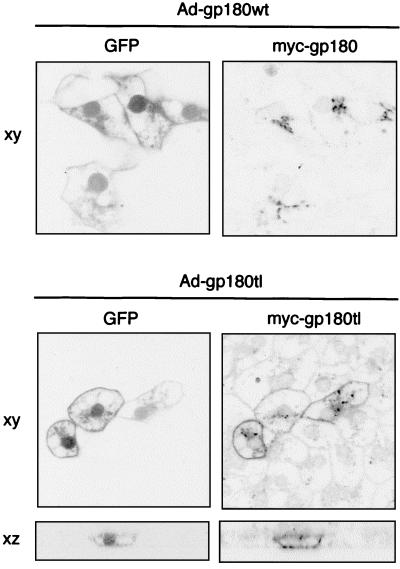
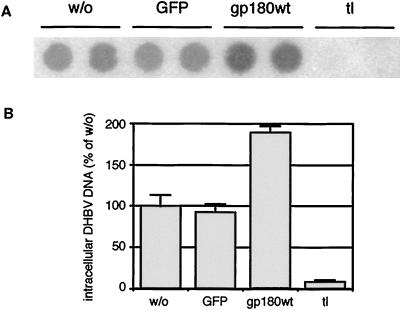
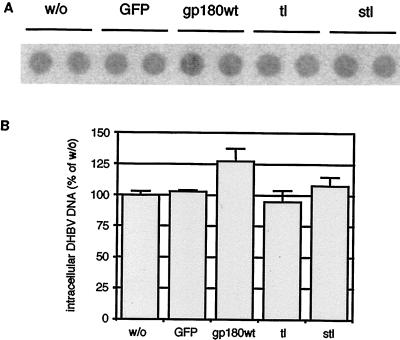
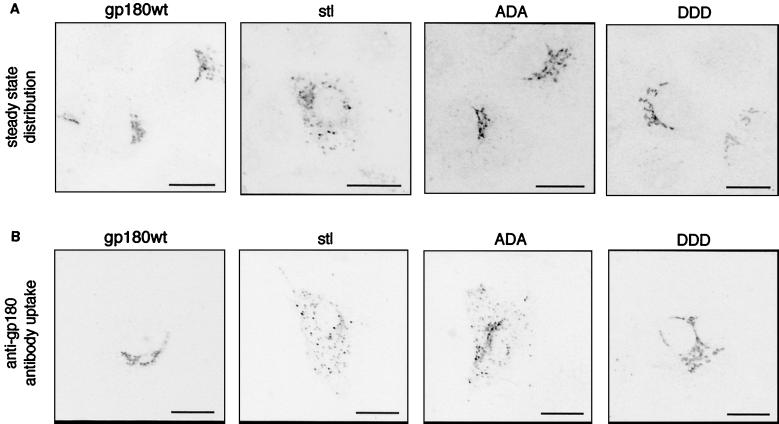
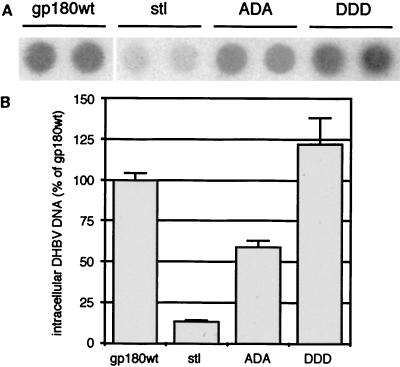
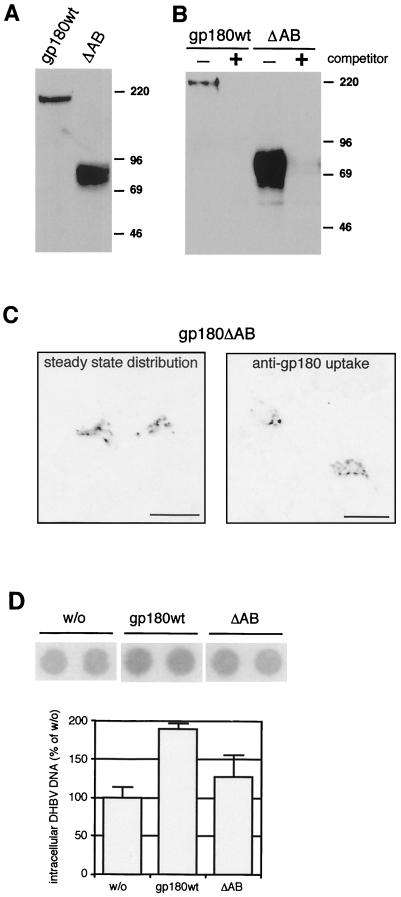
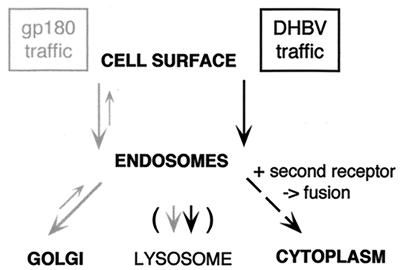
We have investigated the mechanism of duck hepatitis B virus (DHBV) entry into susceptible primary duck hepatocytes (PDHs), using mutants of carboxypeptidase D (gp180), a transmembrane protein shown to act as the primary cellular receptor for avian hepatitis B virus uptake. The variant proteins were abundantly produced from recombinant adenoviruses and tested for the potential to functionally outcompete the endogenous wild-type receptor. Overexpression of wild-type gp180 significantly enhanced the efficiency of DHBV infection in PDHs but did not affect ongoing DHBV replication, an observation further supporting gp180 receptor function. A gp180 mutant deficient for endocytosis abolished DHBV infection, indicating endocytosis to be the route of hepadnaviral entry. With further gp180 variants, carrying mutations in the cytoplasmic domain and characterized by an accelerated turnover, the ability of gp180 to function as a DHBV receptor was found to depend on a wild-type-like sorting phenotype which largely avoids transport toward the endolysosomal compartment. Based on these data, we propose a model in which a distinct intracellular DHBV traffic to the endosome, but not beyond, is a prerequisite for completion of viral entry, i.e., for fusion and capsid release. Furthermore, the deletion of the two enzymatically active carboxypeptidase domains of gp180 did not lead to a loss of receptor function.
Figures
References
-
- Breiner K M. Carboxypeptidase D (gp180): Rezeptor, Transrezeptor und Signalmolekül für Vogelhepatitis B Viren. Ph.D. thesis. Heidelberg, Germany: University of Heidelberg; 1998.
-
- DeMeyer S, Gong J Z, Suwandhi W, van Pelt J, Soumillon A, Yap S H. Organ and species specificity of hepatitis B virus (HBV) infection: a review of literature with a special reference to preferential attachment of HBV to human hepatocytes. J Viral Hepat. 1997;4:145–153. - PubMed
-
- Eng F J, Novikova E G, Kuroki K, Ganem D, Fricker L D. gp180, a protein that binds duck hepatitis B virus particles, has metallocarboxypeptidase D-like enzymatic activity. J Biol Chem. 1998;273:8382–8388. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
