

Relative sensitivity of hepatitis B virus and other hepatotropic viruses to the antiviral effects of cytokines
- PMID: 10666256
- PMCID: PMC111707
- DOI: 10.1128/jvi.74.5.2255-2264.2000
Relative sensitivity of hepatitis B virus and other hepatotropic viruses to the antiviral effects of cytokines
Abstract
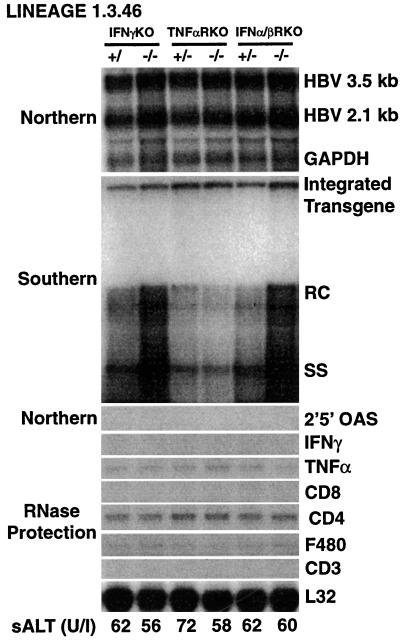
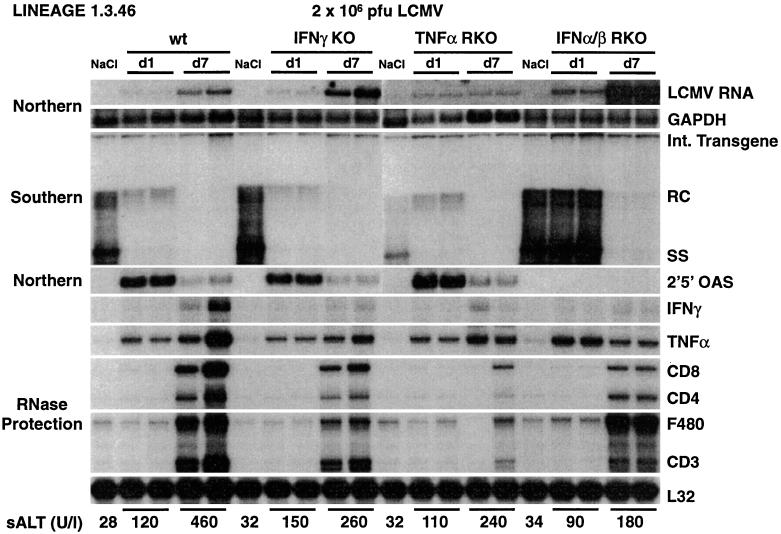
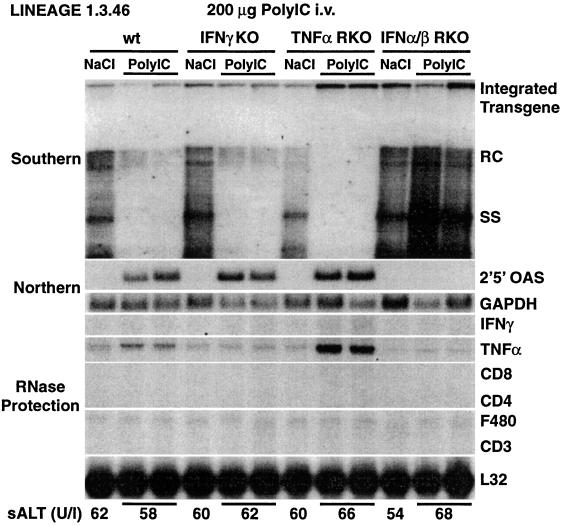
We have previously shown that hepatitis B virus (HBV) replication is inhibited noncytopathically in the livers of transgenic mice following injection of HBV-specific cytotoxic T lymphocytes (CTLs) or infection with unrelated hepatotropic viruses, including lymphocytic choriomeningitis virus (LCMV) and adenovirus. These effects are mediated by gamma interferon (IFNgamma), tumor necrosis factor alpha (TNFalpha), and IFNalpha/beta. In the present study, we crossed HBV transgenic mice with mice genetically deficient for IFNgamma (IFNgammaKO), the TNFalpha receptor (TNFalphaRKO), or the IFNalpha/beta receptor (IFNalpha/betaRKO) in order to determine the relative contribution of each cytokine to the antiviral effects observed in each of these systems. Interestingly, we showed that HBV replicates in unmanipulated IFNgammaKO and IFNalpha/betaRKO mice at levels higher than those observed in control mice, implying that baseline levels of these cytokines control HBV replication in the absence of inflammation. We also showed that IFNgamma mediates most of the antiviral effect of the CTLs while IFNalpha/beta is primarily responsible for the early inhibitory effect of LCMV and adenovirus on HBV replication. In addition, we showed that the hepatic induction of IFNalpha/beta observed after injection of poly(I. C) is sufficient to inhibit HBV replication and that a similar antiviral effect is achieved by systemic administration of very high doses of IFNalpha. We also compared the relative sensitivity of LCMV and adenovirus to control by IFNgamma, TNFalpha, or IFNalpha/beta in these animals. Importantly, IFNalpha/betaRKO mice, and to a lesser extent IFNgammaKO mice, showed higher hepatic levels of LCMV RNA and adenovirus DNA and RNA than control mice, underscoring the importance of both interferons in controlling these other viral infections as well.
Figures




References
-
- Beutler B, Greenwald D, Hulmes J D, Chang M, Pan Y C, Mathison J, Ulevitch R, Cerami A. Identity of tumour necrosis factor and the macrophage-secreted factor cachectin. Nature. 1985;316:552–554. - PubMed
-
- Chisari F V, Ferrari C. Hepatitis B virus immunopathogenesis. Annu Rev Immunol. 1995;13:29–60. - PubMed
-
- Dalton D K, Pitts-Meek S, Keshav S, Figari I S, Bradley A, Stewart T A. Multiple defects of immune cell function in mice with disrupted interferon-γ genes. Science. 1993;259:1739–1742. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
