

Molecular mechanism for the Shp-2 tyrosine phosphatase function in promoting growth factor stimulation of Erk activity
- PMID: 10669730
- PMCID: PMC85329
- DOI: 10.1128/MCB.20.5.1526-1536.2000
Molecular mechanism for the Shp-2 tyrosine phosphatase function in promoting growth factor stimulation of Erk activity
Abstract
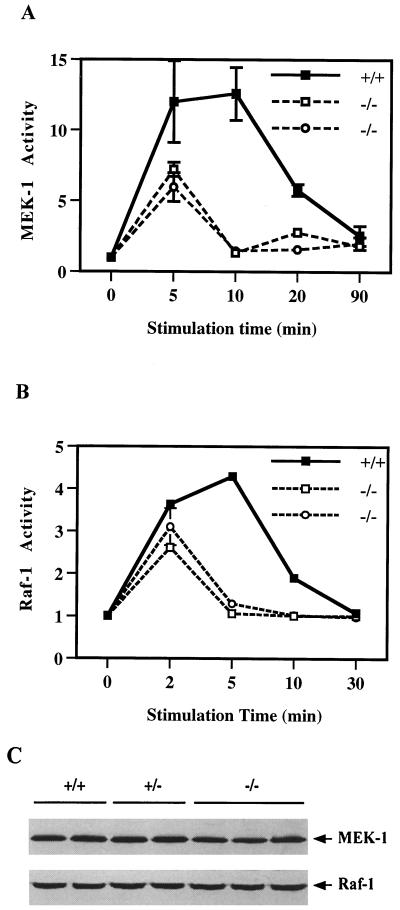
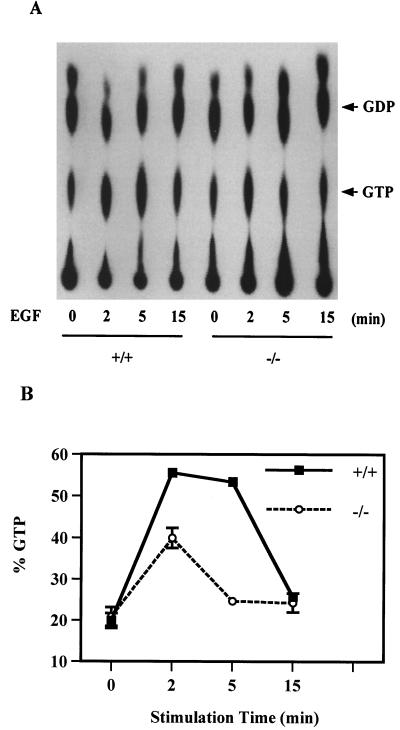
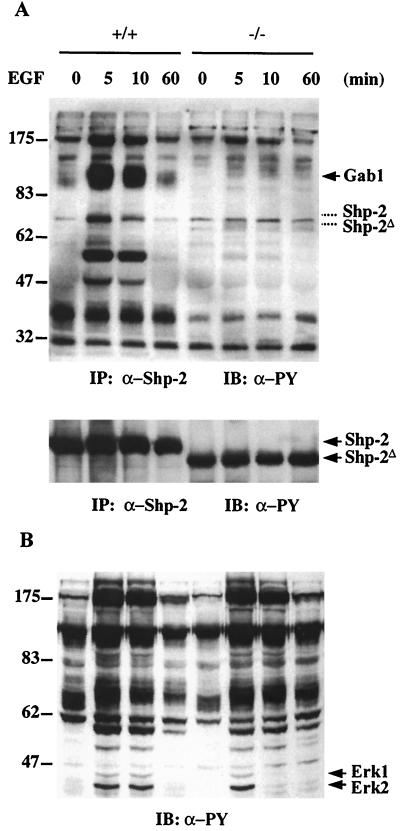
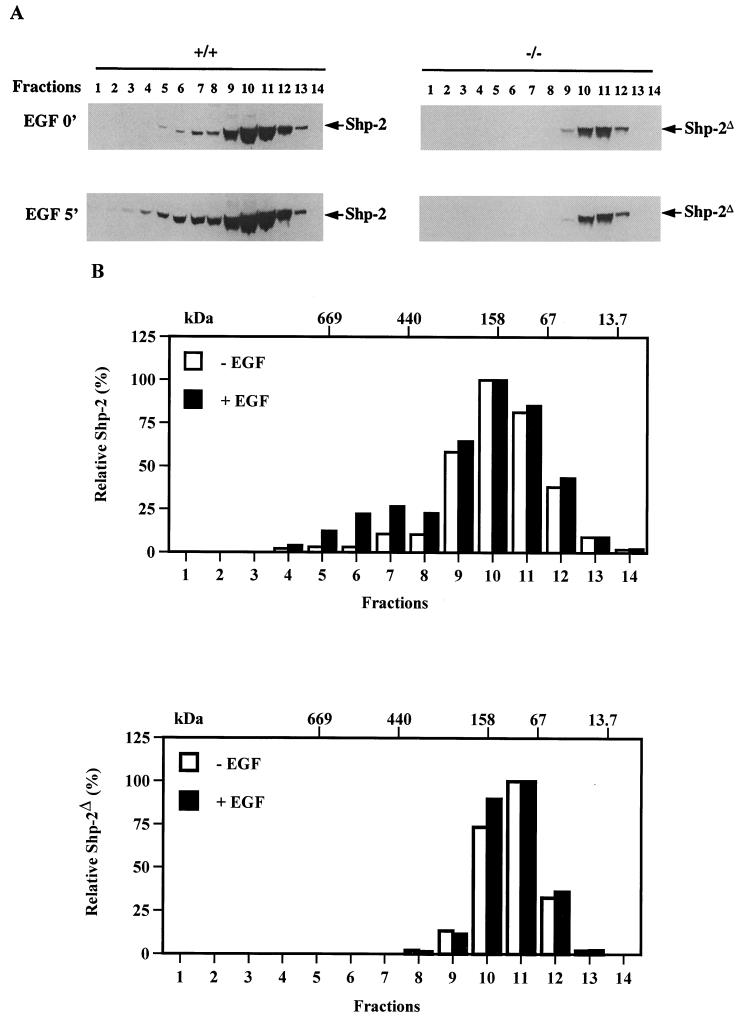
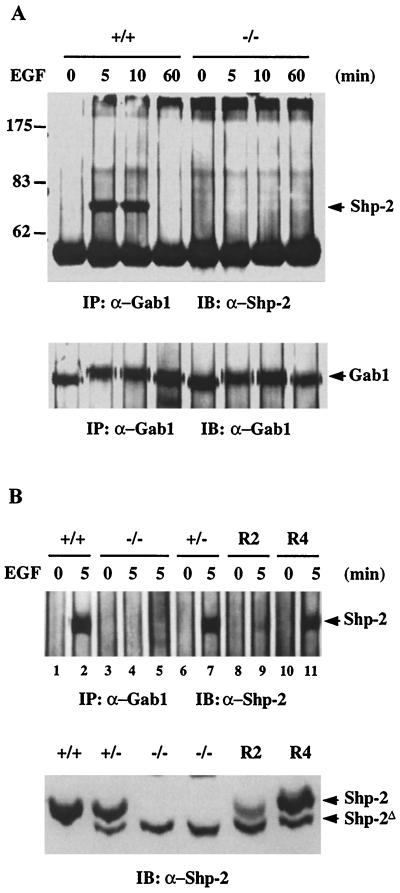
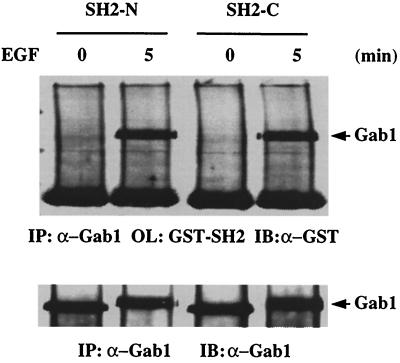
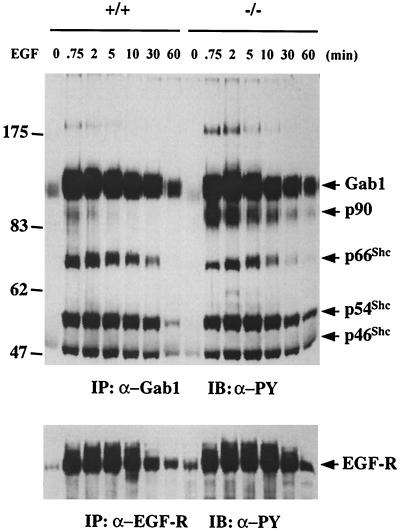
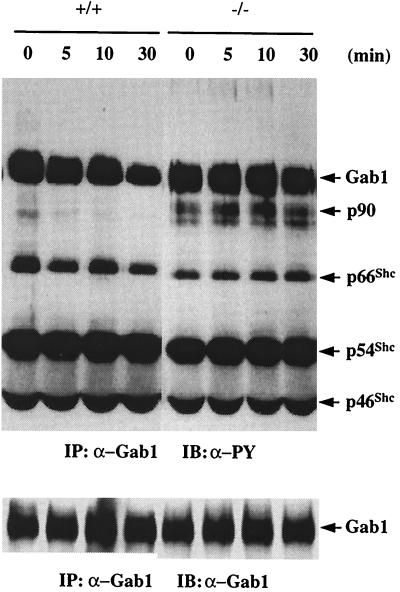
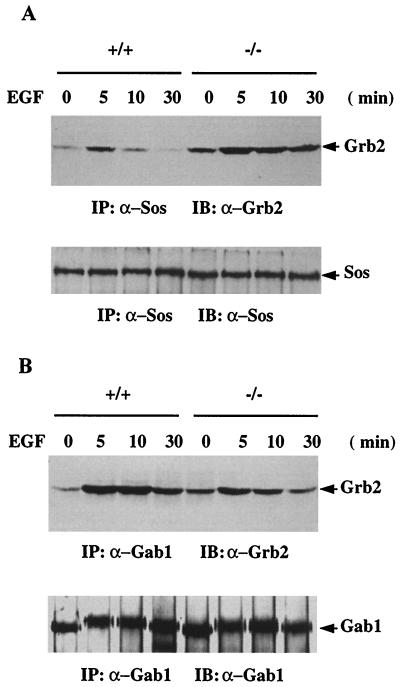
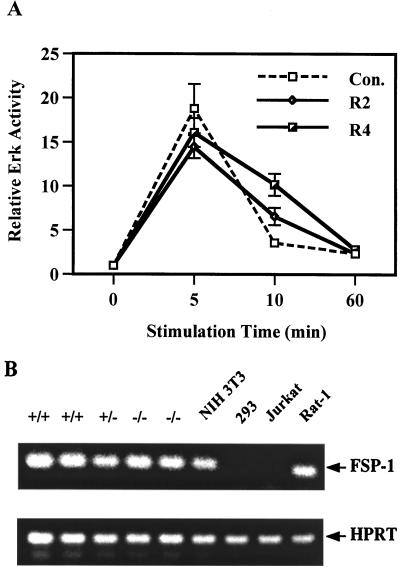
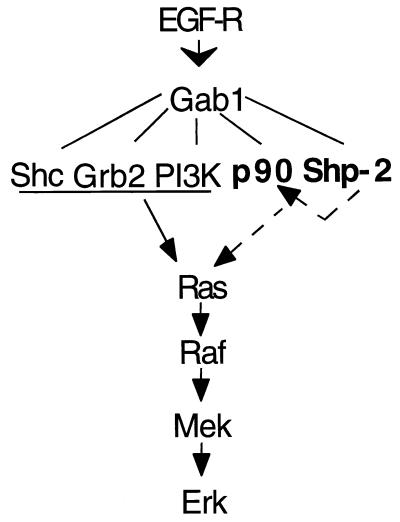
We have previously shown that activation of extracellular signal-regulated kinase (Erk) by epidermal growth factor (EGF) treatment was significantly decreased in mouse fibroblast cells expressing a mutant Shp-2 molecule lacking 65 amino acids in the SH2-N domain, Shp-2(Delta46-110). To address the molecular mechanism for the positive role of Shp-2 in mediating Erk induction, we evaluated the activation of signaling components upstream of Erk in Shp-2 mutant cells. EGF-stimulated Ras, Raf, and Mek activation was significantly attenuated in Shp-2 mutant cells, suggesting that Shp-2 acts to promote Ras activation or to suppress the down-regulation of activated Ras. Biochemical analyses indicate that upon EGF stimulation, Shp-2 is recruited into a multiprotein complex assembled on the Gab1 docking molecule and that Shp-2 seems to exert its biological function by specifically dephosphorylating an unidentified molecule of 90 kDa in the complex. The mutant Shp-2(Delta46-110) molecule failed to participate in the Gab1-organized complex for dephosphorylation of p90, correlating with a defective activation of the Ras-Raf-Mek-Erk cascade in EGF-treated Shp-2 mutant cells. Evidence is also presented that Shp-2 does not appear to modulate the signal relay from EGF receptor to Ras through the Shc, Grb2, and Sos proteins. These results begin to elucidate the mechanism of Shp-2 function downstream of a receptor tyrosine kinase to promote the activation of the Ras-Erk pathway, with potential therapeutic applications in cancer treatment.
Figures
References
-
- Allard J D, Chang H C, Herbst R, McNeill H, Simon M A. The SH2-containing tyrosine phosphatase corkscrew is required during signaling by sevenless, Ras1 and Raf. Development. 1996;122:1137–1146. - PubMed
-
- Barnard D, Diaz B, Clawson D, Marshall M. Oncogenes, growth factors and phorbol esters regulate Raf-1 through common mechanisms. Oncogene. 1998;17:1539–1547. - PubMed
-
- Buday L, Downward J. Epidermal growth factor regulates p21ras through the formation of a complex of receptor, Grb2 adapter protein, and Sos nucleotide exchange factor. Cell. 1993;73:611–620. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous