

Nck-interacting Ste20 kinase couples Eph receptors to c-Jun N-terminal kinase and integrin activation
- PMID: 10669731
- PMCID: PMC85333
- DOI: 10.1128/MCB.20.5.1537-1545.2000
Nck-interacting Ste20 kinase couples Eph receptors to c-Jun N-terminal kinase and integrin activation
Abstract
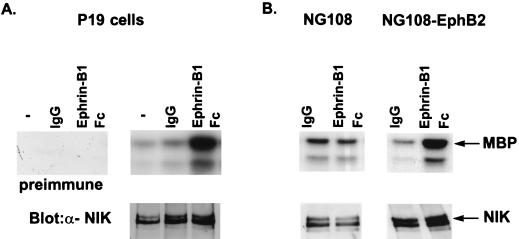
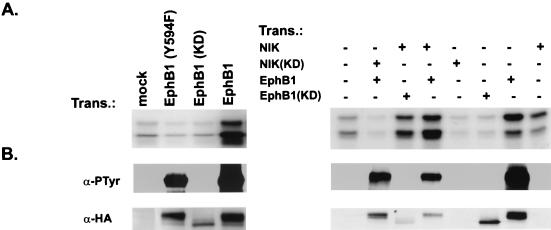
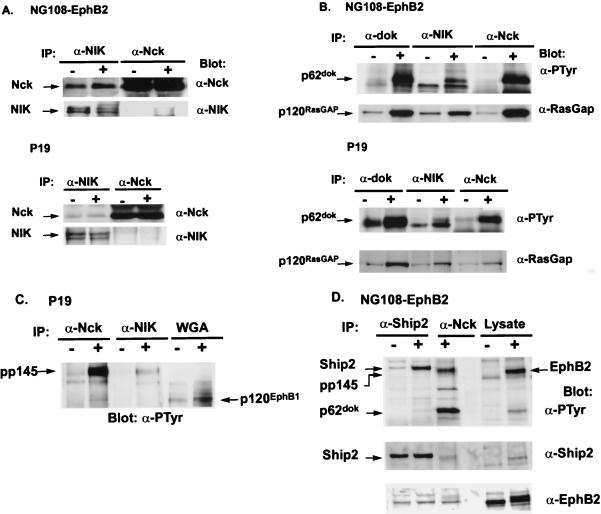
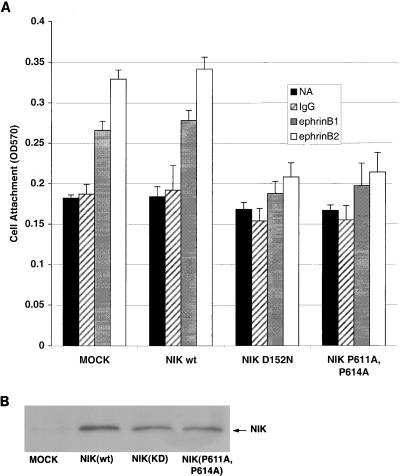
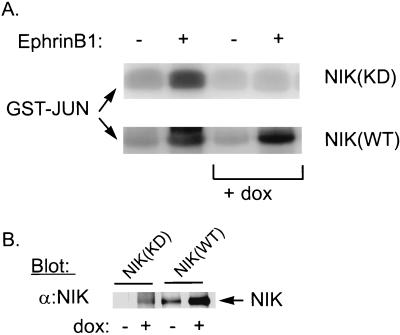
The mammalian Ste20 kinase Nck-interacting kinase (NIK) specifically activates the c-Jun amino-terminal kinase (JNK) mitogen-activated protein kinase module. NIK also binds the SH3 domains of the SH2/SH3 adapter protein Nck. To determine whether Nck functions as an adapter to couple NIK to a receptor tyrosine kinase signaling pathway, we determined whether NIK is activated by Eph receptors (EphR). EphRs constitute the largest family of receptor tyrosine kinases (RTK), and members of this family play important roles in patterning of the nervous and vascular systems. In this report, we show that NIK kinase activity is specifically increased in cells stimulated by two EphRs, EphB1 and EphB2. EphB1 kinase activity and phosphorylation of a juxtamembrane tyrosine (Y594), conserved in all Eph receptors, are both critical for NIK activation by EphB1. Although pY594 in the EphB1R has previously been shown to bind the SH2 domain of Nck, we found that stimulation of EphB1 and EphB2 led predominantly to a complex between NIK/Nck, p62(dok), RasGAP, and an unidentified 145-kDa tyrosine-phosphorylated protein. Tyrosine-phosphorylated p62(dok) most probably binds directly to the SH2 domain of Nck and RasGAP and indirectly to NIK bound to the SH3 domain of Nck. We found that NIK activation is also critical for coupling EphB1R to biological responses that include the activation of integrins and JNK by EphB1. Taken together, these findings support a model in which the recruitment of the Ste20 kinase NIK to phosphotyrosine-containing proteins by Nck is an important proximal step in the signaling cascade downstream of EphRs.
Figures
References
-
- Bohme B, VandenBos T, Cerretti D P, Park L S, Holtrich U, Rubsamen-Waigmann H, Strebhardt K. Cell-cell adhesion mediated by binding of membrane-anchored ligandLERK-2 to the EPH-related receptor human embryonal kinase 2 promotes tyrosine kinase activity. J Biol Chem. 1996;217:24747–24752. - PubMed
-
- Braverman L E, Quilliam L A. Identification of Grb4/Nckbeta, a src homology 2 and 3 domain-containing adapter protein having similar binding and biological properties to Nck. J Biol Chem. 1999;274:5542–5549. - PubMed
-
- Bruckner K, Pasquale E B, Klein R. Tyrosine phosphorylation of transmembrane ligands for Eph receptors. Science. 1997;275:1640–1643. - PubMed
-
- Carpino N, Wisniewski D, Strife A, Marshak D, Kobayashi R, Stillman B, Clarkson B. p62(dok): a constitutively tyrosine-phosphorylated, GAP-associated protein in chronic myelogenous leukemia progenitor cells. Cell. 1997;88:197–204. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous