

Molecular cloning of apobec-1 complementation factor, a novel RNA-binding protein involved in the editing of apolipoprotein B mRNA
- PMID: 10669759
- PMCID: PMC85365
- DOI: 10.1128/MCB.20.5.1846-1854.2000
Molecular cloning of apobec-1 complementation factor, a novel RNA-binding protein involved in the editing of apolipoprotein B mRNA
Abstract
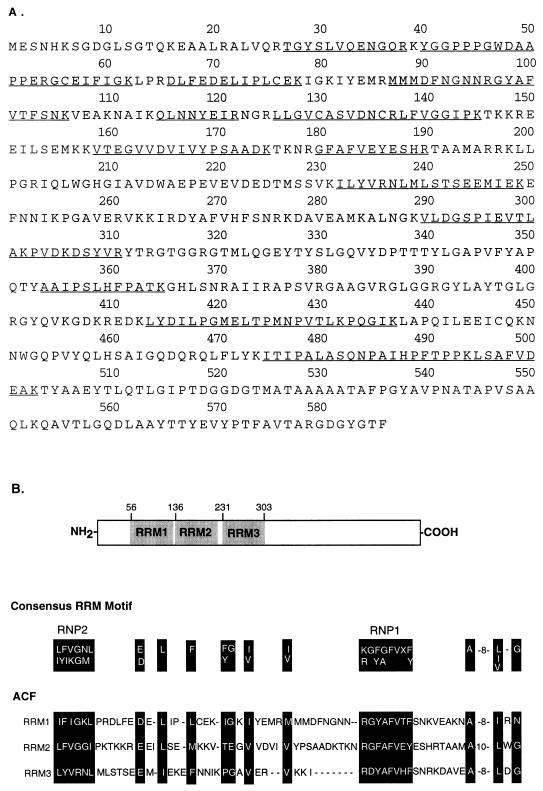
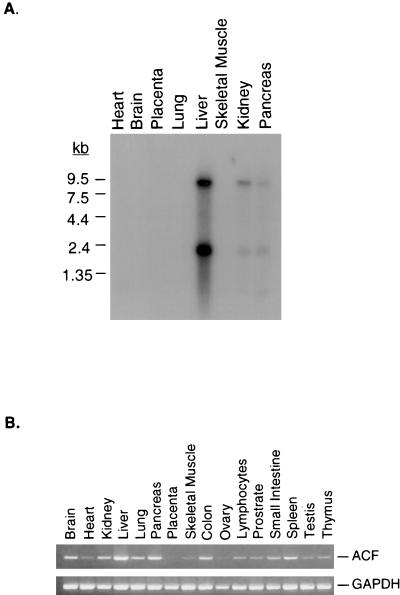
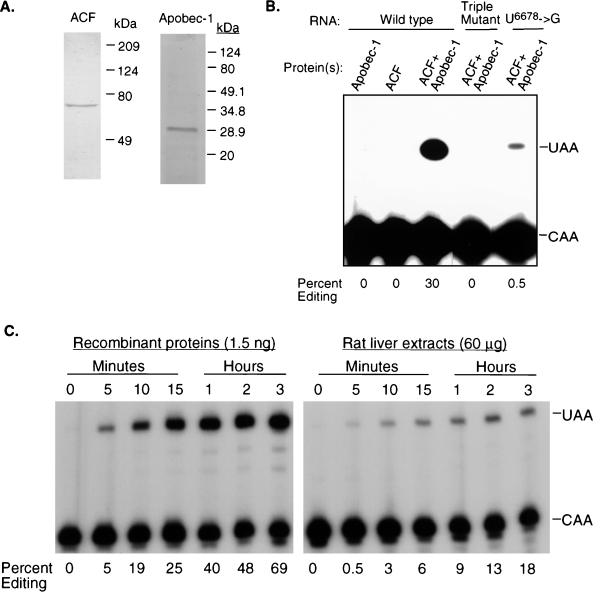
The C-to-U editing of apolipoprotein B (apo-B) mRNA is catalyzed by a multiprotein complex that recognizes an 11-nucleotide mooring sequence downstream of the editing site. The catalytic subunit of the editing enzyme, apobec-1, has cytidine deaminase activity but requires additional unidentified proteins to edit apo-B mRNA. We purified a 65-kDa protein that functionally complements apobec-1 and obtained peptide sequence information which was used in molecular cloning experiments. The apobec-1 complementation factor (ACF) cDNA encodes a novel 64.3-kDa protein that contains three nonidentical RNA recognition motifs. ACF and apobec-1 comprise the minimal protein requirements for apo-B mRNA editing in vitro. By UV cross-linking and immunoprecipitation, we show that ACF binds to apo-B mRNA in vitro and in vivo. Cross-linking of ACF is not competed by RNAs with mutations in the mooring sequence. Coimmunoprecipitation experiments identified an ACF-apobec-1 complex in transfected cells. Immunodepletion of ACF from rat liver extracts abolished editing activity. The immunoprecipitated complexes contained a functional holoenzyme. Our results support a model of the editing enzyme in which ACF binds to the mooring sequence in apo-B mRNA and docks apobec-1 to deaminate its target cytidine. The fact that ACF is widely expressed in human tissues that lack apobec-1 and apo-B mRNA suggests that ACF may be involved in other RNA editing or RNA processing events.
Figures




References
-
- Anant S, Giannoni F, Antic D, DeMaria C T, Keene J D, Brewer G, Davidson N O. AU-rich RNA binding proteins Hel-N1 and AUF1 bind apolipoprotein B mRNA and inhibit posttranscriptional C to U editing. Nucleic Acids Symp Ser. 1997;36:115–118.
-
- Anant S, MacGinnitie A J, Davidson N O. apobec-1, the catalytic subunit of the mammalian apolipoprotein B mRNA editing enzyme, is a novel RNA-binding protein. J Biol Chem. 1995;270:14762–14767. - PubMed
-
- Bass B L. RNA editing and hypermutation by adenosine deamination. Trends Biochem Sci. 1997;22:157–162. . (Erratum, 22:278.) - PubMed
-
- Burd C G, Dreyfuss G. Conserved structures and diversity of functions of RNA-binding proteins. Science. 1994;265:615–621. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous