

The BIR motifs mediate dominant interference and oligomerization of inhibitor of apoptosis Op-IAP
- PMID: 10669762
- PMCID: PMC85372
- DOI: 10.1128/MCB.20.5.1877-1885.2000
The BIR motifs mediate dominant interference and oligomerization of inhibitor of apoptosis Op-IAP
Abstract
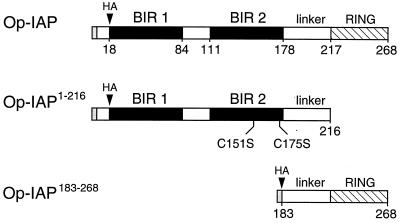
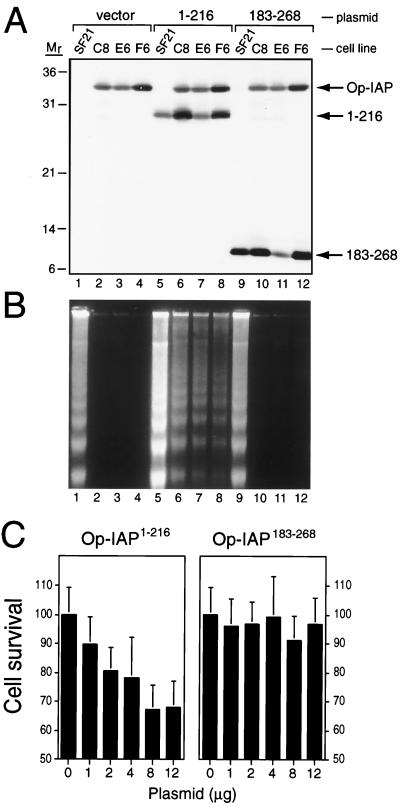
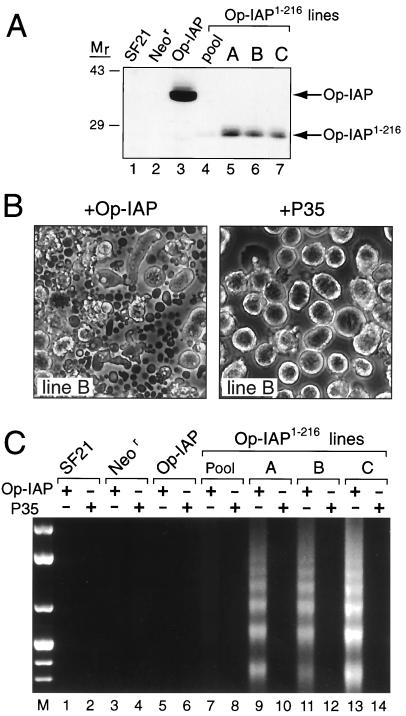
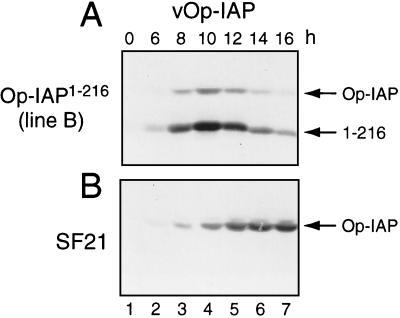
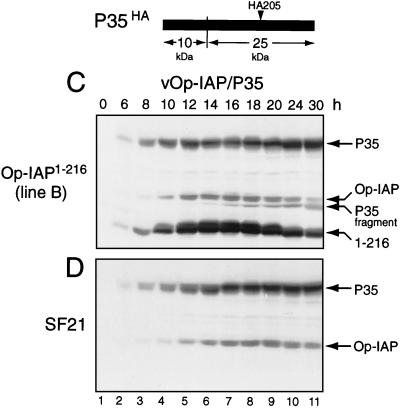
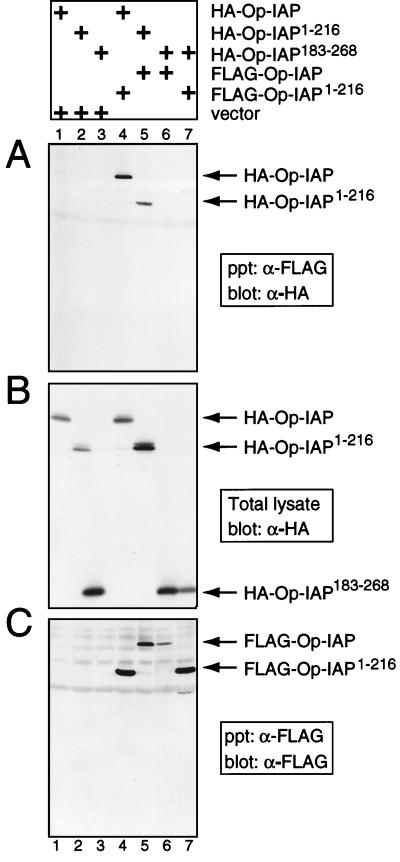
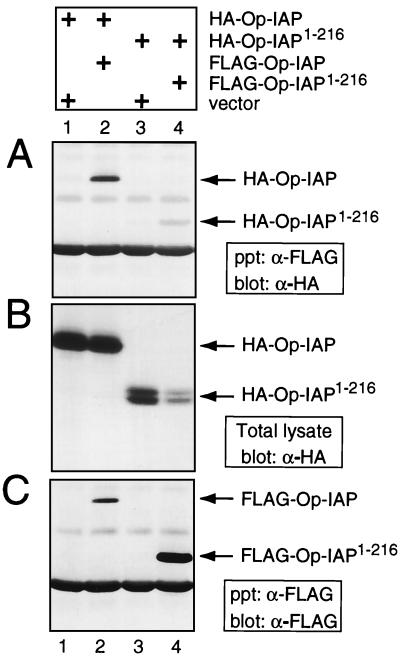
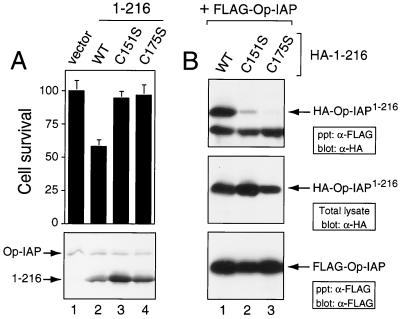
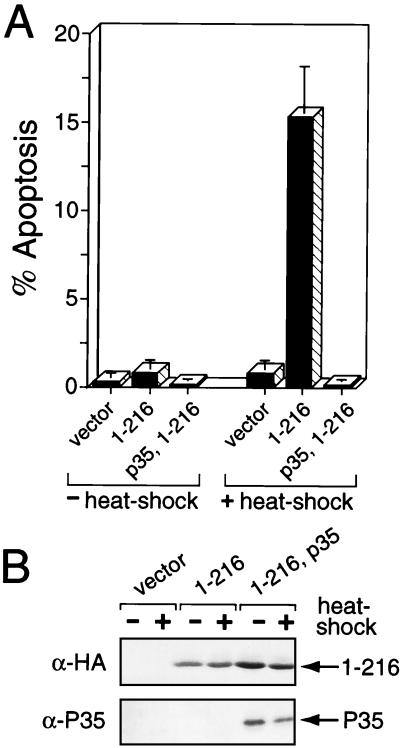
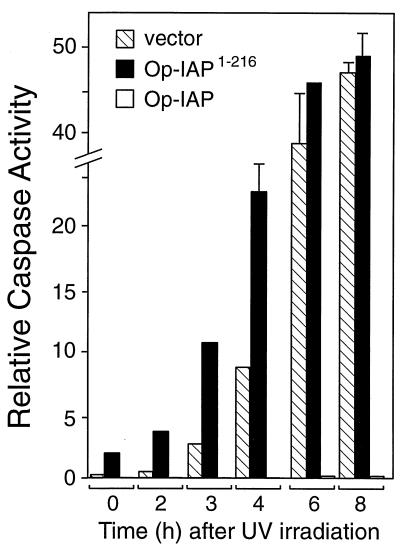
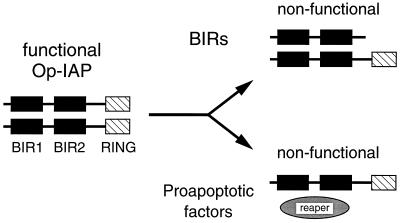
The defining structural motif of the inhibitor of apoptosis (iap) protein family is the BIR (baculovirus iap repeat), a highly conserved zinc coordination domain of approximately 70 residues. Although the BIR is required for inhibitor-of-apoptosis (IAP) function, including caspase inhibition, its molecular role in antiapoptotic activity in vivo is unknown. To define the function of the BIRs, we investigated the activity of these structural motifs within Op-IAP, an efficient, virus-derived IAP. We report here that Op-IAP(1-216), a loss-of-function truncation which contains two BIRs but lacks the C-terminal RING motif, potently interfered with Op-IAP's capacity to block apoptosis induced by diverse stimuli. In contrast, Op-IAP(1-216) had no effect on apoptotic suppression by caspase inhibitor P35. Consistent with a mechanism of dominant inhibition that involves direct interaction between Op-IAP(1-216) and full-length Op-IAP, both proteins formed an immunoprecipitable complex in vivo. Op-IAP also self-associated. In contrast, the RING motif-containing truncation Op-IAP(183-268) failed to interact with or interfere with Op-IAP function. Substitution of conserved residues within BIR 2 caused loss of dominant inhibition by Op-IAP(1-216) and coincided with loss of interaction with Op-IAP. Thus, residues encompassing the BIRs mediate dominant inhibition and oligomerization of Op-IAP. Consistent with dominant interference by interaction with an endogenous cellular IAP, Op-IAP(1-216) also lowered the survival threshold of cultured insect cells. Taken together, these data suggest a new model wherein the antiapoptotic function of IAP requires homo-oligomerization, which in turn mediates specific interactions with cellular apoptotic effectors.
Figures
Similar articles
-
A mutational analysis of the baculovirus inhibitor of apoptosis Op-IAP.J Biol Chem. 1998 Dec 18;273(51):33915-21. doi: 10.1074/jbc.273.51.33915. J Biol Chem. 1998. PMID: 9852042
-
Baculovirus Inhibitor-of-Apoptosis Op-IAP3 Blocks Apoptosis by Interaction with and Stabilization of a Host Insect Cellular IAP.J Virol. 2015 Oct 21;90(1):533-44. doi: 10.1128/JVI.02320-15. Print 2016 Jan 1. J Virol. 2015. PMID: 26491164 Free PMC article.
-
Baculovirus inhibitor of apoptosis functions at or upstream of the apoptotic suppressor P35 to prevent programmed cell death.J Virol. 1997 Jun;71(6):4509-16. doi: 10.1128/JVI.71.6.4509-4516.1997. J Virol. 1997. PMID: 9151843 Free PMC article.
-
An exegesis of IAPs: salvation and surprises from BIR motifs.Trends Cell Biol. 1999 Aug;9(8):323-8. doi: 10.1016/s0962-8924(99)01609-8. Trends Cell Biol. 1999. PMID: 10407412 Review.
-
Targeting the BIR Domains of Inhibitor of Apoptosis (IAP) Proteins in Cancer Treatment.Comput Struct Biotechnol J. 2019 Jan 25;17:142-150. doi: 10.1016/j.csbj.2019.01.009. eCollection 2019. Comput Struct Biotechnol J. 2019. PMID: 30766663 Free PMC article. Review.
Cited by
-
Molecular Pathogenesis of MALT Lymphoma.Gastroenterol Res Pract. 2015;2015:102656. doi: 10.1155/2015/102656. Epub 2015 Apr 1. Gastroenterol Res Pract. 2015. PMID: 25922601 Free PMC article. Review.
-
Host insect inhibitor-of-apoptosis SfIAP functionally replaces baculovirus IAP but is differentially regulated by Its N-terminal leader.J Virol. 2010 Nov;84(21):11448-60. doi: 10.1128/JVI.01311-10. Epub 2010 Aug 25. J Virol. 2010. PMID: 20739517 Free PMC article.
-
Identification and functional analysis of the putative anti-apoptotic gene iap4 of Spodoptera litura nucleopolyhedrovirus.Virus Genes. 2010 Oct;41(2):298-304. doi: 10.1007/s11262-010-0513-4. Epub 2010 Jul 13. Virus Genes. 2010. PMID: 20625923
-
Insect inhibitor-of-apoptosis (IAP) proteins are negatively regulated by signal-induced N-terminal degrons absent within viral IAP proteins.J Virol. 2015 Apr;89(8):4481-93. doi: 10.1128/JVI.03659-14. Epub 2015 Feb 4. J Virol. 2015. PMID: 25653450 Free PMC article.
-
Heliothis zea nudivirus 1 gene hhi1 induces apoptosis which is blocked by the Hz-iap2 gene and a noncoding gene, pag1.J Virol. 2011 Jul;85(14):6856-66. doi: 10.1128/JVI.01843-10. Epub 2011 May 4. J Virol. 2011. PMID: 21543471 Free PMC article.
References
-
- Adida C, Berrebi D, Peuchmaur M, Reyes-Mugica M, Altieri D C. Anti-apoptotis gene, survivin, and prognosis of neuroblastoma. Lancet. 1998;351:882–883. - PubMed
-
- Ambrosini G, Adida C, Altieri D C. A novel anti-apoptosis gene, survivin, expressed in cancer and lymphoma. Nat Med. 1997;3:917–921. - PubMed
-
- Bergmann A, Agapite J, Steller H. Mechanisms and control of programmed cell death in invertebrates. Oncogene. 1998;17:3215–3223. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous