

Regulation of cold shock-induced RNA helicase gene expression in the Cyanobacterium anabaena sp. strain PCC 7120
- PMID: 10671444
- PMCID: PMC94409
- DOI: 10.1128/JB.182.5.1251-1256.2000
Regulation of cold shock-induced RNA helicase gene expression in the Cyanobacterium anabaena sp. strain PCC 7120
Abstract
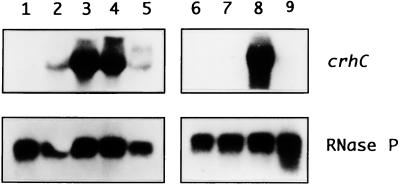
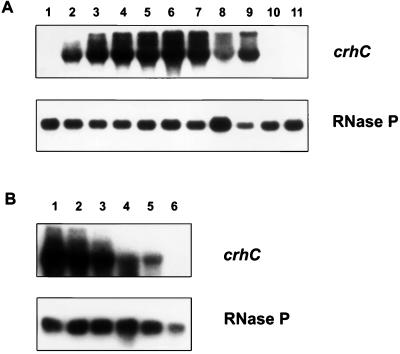
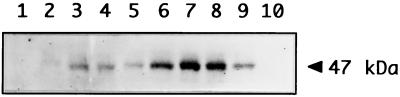
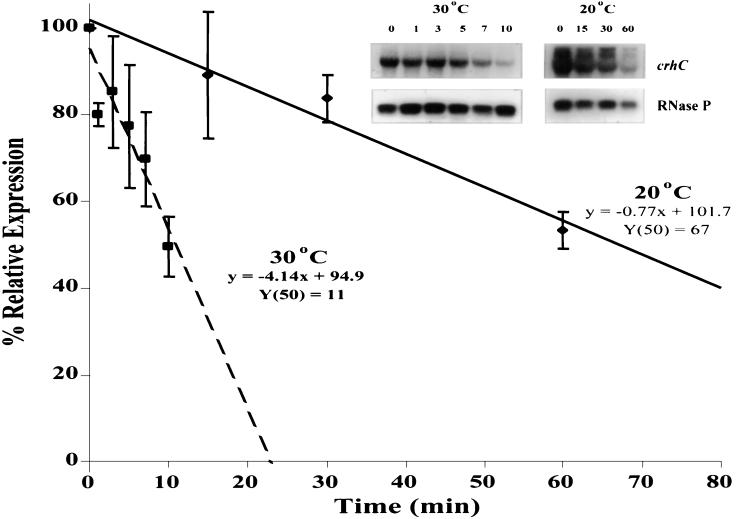
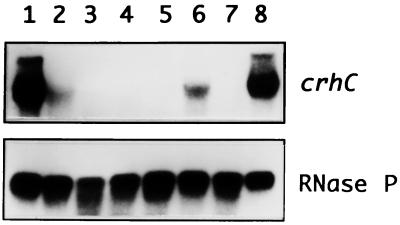
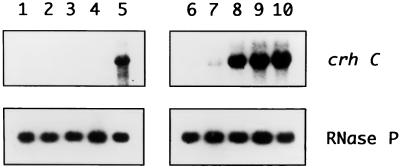
Expression of the Anabaena sp. strain PCC 7120 RNA helicase gene crhC is induced by cold shock. crhC transcripts are not detectable at 30 degrees C but accumulate at 20 degrees C, and levels remain elevated for the duration of the cold stress. Light-derived metabolic capability, and not light per se, is required for crhC transcript accumulation. Enhanced crhC mRNA stability contributes significantly to the accumulation of crhC transcripts, with the crhC half-life increasing sixfold at 20 degrees C. The accumulation is reversible, with the cells responding more rapidly to temperature downshifts than to upshifts, as a result of the lack of active mRNA destabilization and the continuation of crhC transcription, at least transiently, after a temperature upshift. Translational inhibitors do not induce crhC expression to cold shock levels, indicating that inhibition of translation is only one of the signals required to activate the cold shock response in Anabaena. Limited amounts of protein synthesis are required for the cold shock-induced accumulation of crhC transcripts, as normal levels of accumulation occur in the presence of tetracycline but are abolished by chloramphenicol. Regulation of crhC expression may also extend to the translational level, as CrhC protein levels do not correlate completely with the pattern of mRNA transcript accumulation. Our experiments indicate that the regulation of crhC transcript accumulation is tightly controlled by both temperature and metabolic activity at the levels of transcription, mRNA stabilization, and translation.
Figures
Similar articles
-
Characterization of the cold stress-induced cyanobacterial DEAD-box protein CrhC as an RNA helicase.Nucleic Acids Res. 2000 Oct 15;28(20):3926-34. doi: 10.1093/nar/28.20.3926. Nucleic Acids Res. 2000. PMID: 11024172 Free PMC article.
-
Polar-biased localization of the cold stress-induced RNA helicase, CrhC, in the Cyanobacterium Anabaena sp. strain PCC 7120.Mol Microbiol. 2003 Nov;50(4):1439-48. doi: 10.1046/j.1365-2958.2003.03783.x. Mol Microbiol. 2003. PMID: 14622428
-
A cold shock-induced cyanobacterial RNA helicase.J Bacteriol. 1999 Mar;181(6):1728-32. doi: 10.1128/JB.181.6.1728-1732.1999. J Bacteriol. 1999. PMID: 10074063 Free PMC article.
-
RNA remodeling and gene regulation by cold shock proteins.RNA Biol. 2010 Nov-Dec;7(6):788-95. doi: 10.4161/rna.7.6.13482. Epub 2010 Nov 1. RNA Biol. 2010. PMID: 21045540 Free PMC article. Review.
-
Transcriptional and post-transcriptional control of cold-shock genes.J Mol Biol. 2003 Aug 15;331(3):527-39. doi: 10.1016/s0022-2836(03)00732-0. J Mol Biol. 2003. PMID: 12899826 Review.
Cited by
-
Autoregulation of RNA helicase expression in response to temperature stress in Synechocystis sp. PCC 6803.PLoS One. 2012;7(10):e48683. doi: 10.1371/journal.pone.0048683. Epub 2012 Oct 31. PLoS One. 2012. PMID: 23119089 Free PMC article.
-
Liquid-Liquid Phase Separation of the DEAD-Box Cyanobacterial RNA Helicase Redox (CrhR) into Dynamic Membraneless Organelles in Synechocystis sp. Strain PCC 6803.Appl Environ Microbiol. 2023 Apr 26;89(4):e0001523. doi: 10.1128/aem.00015-23. Epub 2023 Mar 15. Appl Environ Microbiol. 2023. PMID: 36920190 Free PMC article.
-
A LexA-related protein regulates redox-sensitive expression of the cyanobacterial RNA helicase, crhR.Nucleic Acids Res. 2006 Jul 13;34(12):3446-54. doi: 10.1093/nar/gkl426. Print 2006. Nucleic Acids Res. 2006. PMID: 16840531 Free PMC article.
-
Characterization of the cold stress-induced cyanobacterial DEAD-box protein CrhC as an RNA helicase.Nucleic Acids Res. 2000 Oct 15;28(20):3926-34. doi: 10.1093/nar/28.20.3926. Nucleic Acids Res. 2000. PMID: 11024172 Free PMC article.
-
Effects of low temperature on tropical and temperate isolates of marine Synechococcus.ISME J. 2016 May;10(5):1252-63. doi: 10.1038/ismej.2015.179. Epub 2015 Oct 23. ISME J. 2016. PMID: 26495993 Free PMC article.
References
-
- Ausubel F M, Brent R, Kingston R E, Moore D D, Seidman J G, Smith J A, Struhl K. Current protocols in molecular biology. New York, N.Y: John Wiley & Sons, Inc.; 1994.
-
- Brandi A, Pietroni P, Gualerzi C O, Pon C L. Post-transcriptional regulation of CspA expression in Escherichia coli. Mol Microbiol. 1996;19:231–240. - PubMed
-
- Fang L, Jiang W, Bae W, Inouye M. Promoter-independent cold-shock induction of cspA and its derepression at 37°C by mRNA stabilization. Mol Microbiol. 1997;23:355–364. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
