

Voltage-gated currents distinguish parvocellular from magnocellular neurones in the rat hypothalamic paraventricular nucleus
- PMID: 10673555
- PMCID: PMC2269788
- DOI: 10.1111/j.1469-7793.2000.t01-1-00193.x
Voltage-gated currents distinguish parvocellular from magnocellular neurones in the rat hypothalamic paraventricular nucleus
Abstract
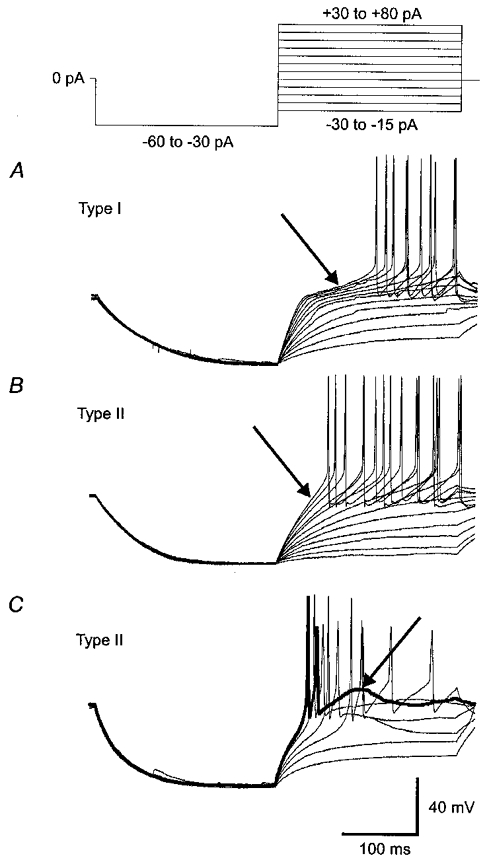
1. Magnocellular and parvocellular neurones of the hypothalamic paraventricular nucleus (PVN) differentially regulate pituitary hormone secretion and autonomic output. Previous experiments have suggested that magnocellular, or type I neurones, and parvocellular, or type II neurones, of the PVN express different electrophysiological properties. Whole-cell patch-clamp recordings were performed in hypothalamic slices to identify the voltage-gated currents responsible for the electrophysiological differences between type I and type II PVN neurones. 2. Type I neurones, which display transient outward rectification and lack a low-threshold spike (LTS), generated a large A-type K+ current (IA) (mean +/- s.e. m.: 1127.5 +/- 126.4 pA; range: 250-3600 pA; voltage steps to -25 mV) but expressed little or no T-type Ca2+ current (IT). Type II neurones, which lack transient outward rectification but often display an LTS, expressed a smaller IA (360.1 +/- 56.3 pA; range: 40-1100 pA; voltage steps to -25 mV), and 75 % of the type II neurones generated an IT (-402.5 +/- 166.9 pA; range: -90 to -2200 pA; at peak). 3. The voltage dependence of IA was shifted to more negative values in type I neurones compared to type II neurones. Thus, the activation threshold (-53.5 +/- 0.9 and -46.1 +/- 2.6 mV), the half-activation potential (-25 +/- 1.9 and -17.9 +/- 2.0 mV), the half-inactivation potential (-80.4 +/- 9.3 and -67.2 +/- 3.0 mV), and the potential at which the current became fully inactivated (-57.4 +/- 2.1 and -49.8 +/- 1.5 mV) were more negative in type I neurones than in type II neurones, respectively. 4. IT in type II neurones activated at a threshold of -59.2 +/- 1.2 mV, peaked at -32. 6 +/- 1.7 mV, was half-inactivated at -66.9 +/- 2.2 mV, and was fully inactivated at -52.2 +/- 2.2 mV. 5. Both cell types expressed a delayed rectifier current with similar voltage dependence, although it was smaller in type I neurones (389.7 +/- 39.3 pA) than in type II neurones (586.4 +/- 76.0 pA). 6. In type I neurones IA was reduced by 41.1 +/- 7.0 % and the action potential delay caused by the transient outward rectification was reduced by 46.2 +/- 10.3 % in 5 mM 4-aminopyridine. In type II neurones IT was reduced by 66.8 +/- 10.9 % and the LTS was reduced by 76.7 +/- 7.8 % in 100 microM nickel chloride, but neither IT nor LTS was sensitive to 50 microM cadmium chloride. 7. Thus, differences in the electrophysiological properties between type I, putative magnocellular neurones and type II, putative parvocellular neurones of the PVN can be attributed to the differential expression of voltage-gated K+ and Ca2+ currents. This diversity of ion channel expression is likely to have profound effects on the response properties of these neurosecretory and non-neurosecretory neurones.
Figures










References
-
- Andreasen M, Hablitz JJ. Kinetic properties of a transient outward current in rat neocortical neurons. Journal of Neurophysiology. 1992;68:1133–1142. - PubMed
-
- Bicknell RJ, Leng G. Relative efficiency of neural firing patterns for vasopressin release in vitro. Neuroendocrinology. 1981;33:295–299. - PubMed
-
- Carbone E, Lux HD. A low voltage-activated, fully inactivating Ca channel in vertebrate sensory neurones. Nature. 1984;310:501–502. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous