

YidC, the Escherichia coli homologue of mitochondrial Oxa1p, is a component of the Sec translocase
- PMID: 10675323
- PMCID: PMC305592
- DOI: 10.1093/emboj/19.4.542
YidC, the Escherichia coli homologue of mitochondrial Oxa1p, is a component of the Sec translocase
Abstract
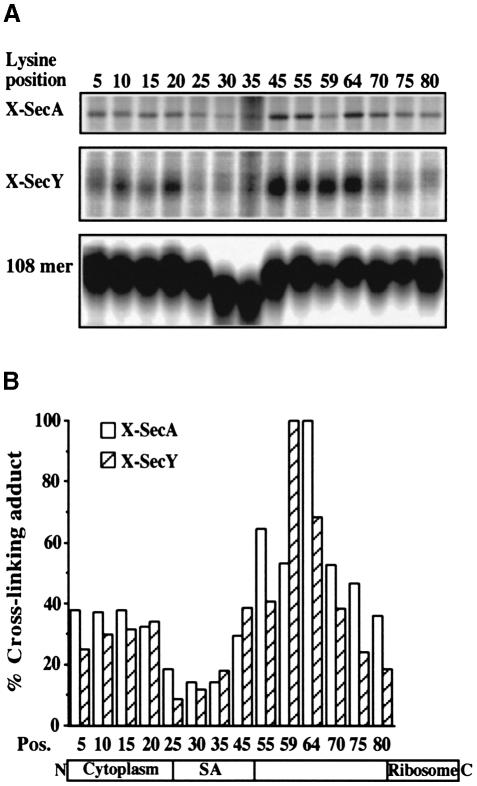
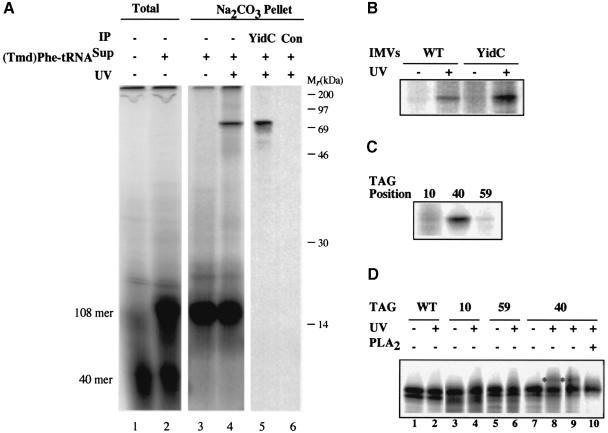
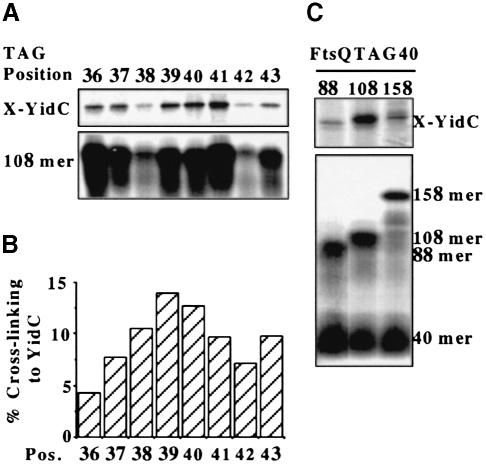
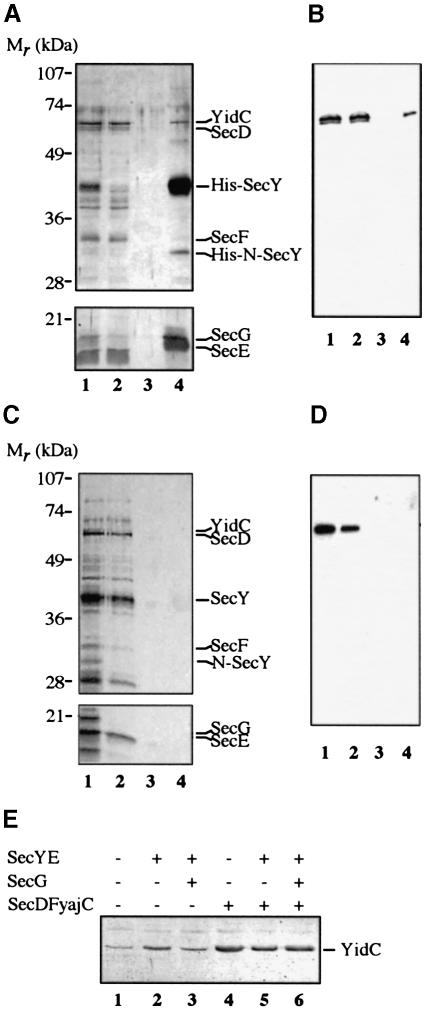
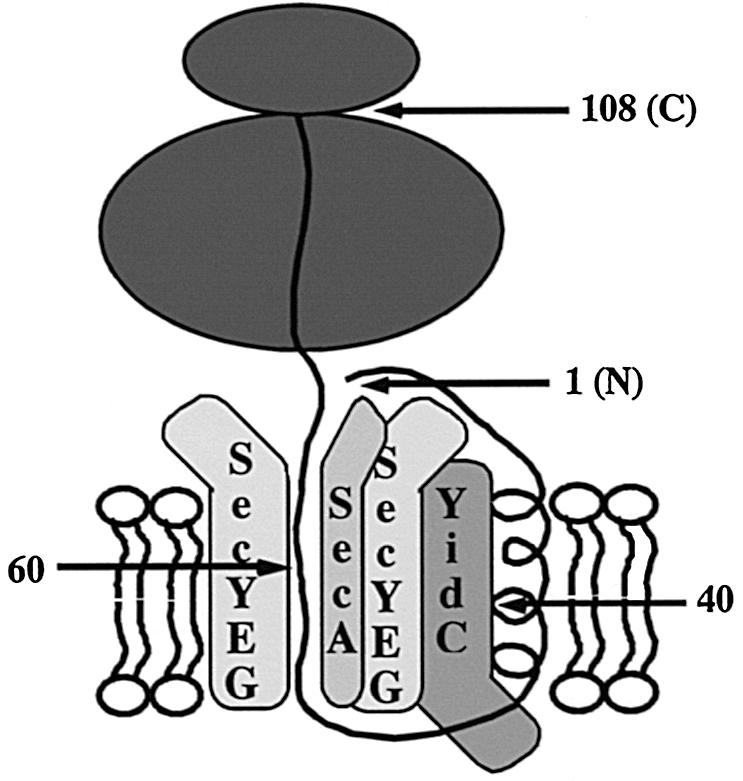
In Escherichia coli, both secretory and inner membrane proteins initially are targeted to the core SecYEG inner membrane translocase. Previous work has also identified the peripherally associated SecA protein as well as the SecD, SecF and YajC inner membrane proteins as components of the translocase. Here, we use a cross-linking approach to show that hydrophilic portions of a co-translationally targeted inner membrane protein (FtsQ) are close to SecA and SecY, suggesting that insertion takes place at the SecA/Y interface. The hydrophobic FtsQ signal anchor sequence contacts both lipids and a novel 60 kDa translocase-associated component that we identify as YidC. YidC is homologous to Saccharomyces cerevisiae Oxa1p, which has been shown to function in a novel export pathway at the mitochondrial inner membrane. We propose that YidC is involved in the insertion of hydrophobic sequences into the lipid bilayer after initial recognition by the SecAYEG translocase.
Figures
References
-
- Brunner J. (1996) Use of photocrosslinkers in cell biology. Trends Cell Biol., 6, 154–157. - PubMed
-
- Cagnon C., Valverde, V. and Masson, J.-M. (1991) A new family of sugar-inducible expression vectors for Escherichia coli.Protein Eng., 4, 843–847. - PubMed
-
- De Gier J.W.L., Mansournia, P., Valent, Q.A., Phillips, G.J., Luirink, J. and von Heijne, G. (1996) Assembly of a cytoplasmic membrane protein in Escherichia coli is dependent on the signal recognition particle. FEBS Lett., 399, 307–309. - PubMed
-
- De Vrije T., Tommassen, J. and de Kruijff, B. (1987) Optimal posttranslational translocation of the precursor of PhoE protein across Escherichia coli membrane vesicles requires both ATP and the proton motive force. Biochim. Biophys. Acta, 900, 63–72. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
