

Op18/stathmin caps a kinked protofilament-like tubulin tetramer
- PMID: 10675326
- PMCID: PMC305595
- DOI: 10.1093/emboj/19.4.572
Op18/stathmin caps a kinked protofilament-like tubulin tetramer
Abstract
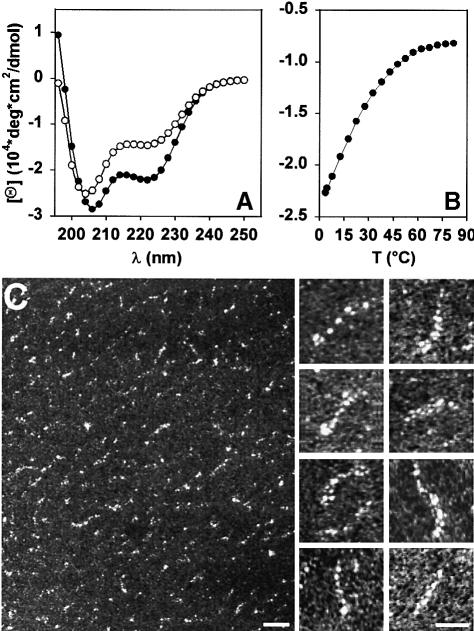
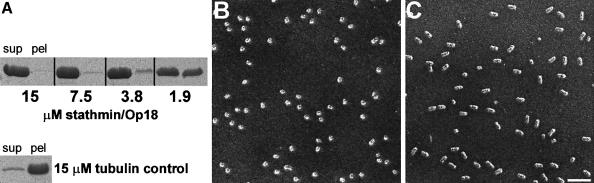
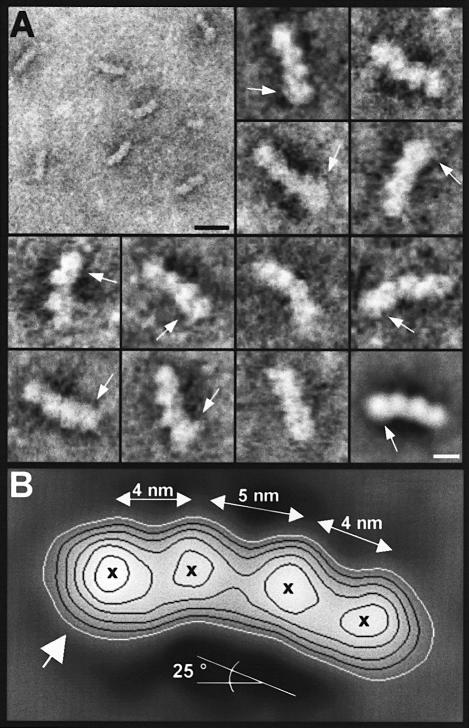
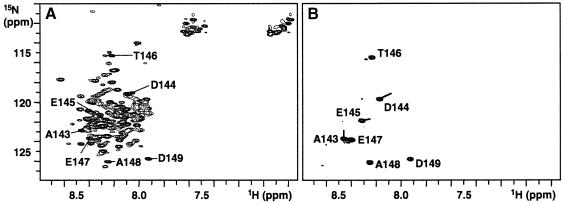
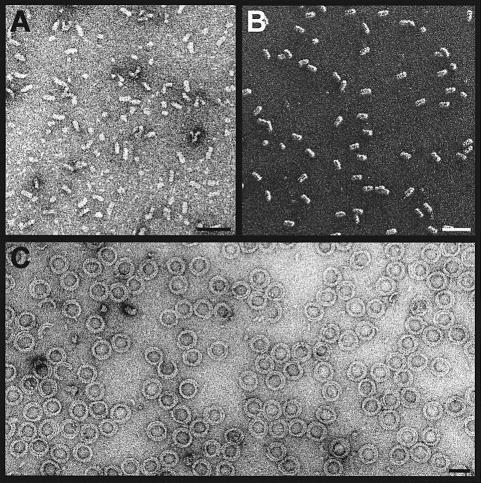
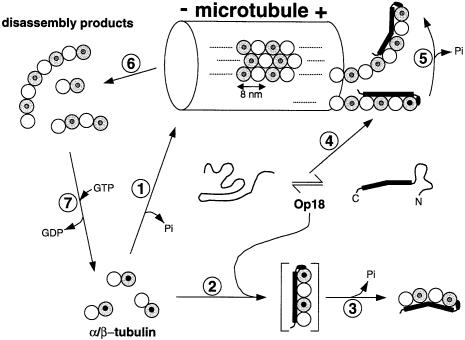
Oncoprotein 18/stathmin (Op18), a regulator of microtubule dynamics, was recombinantly expressed and its structure and function analysed. We report that Op18 by itself can fold into a flexible and extended alpha-helix, which is in equilibrium with a less ordered structure. In complex with tubulin, however, all except the last seven C-terminal residues of Op18 are tightly bound to tubulin. Digital image analysis of Op18:tubulin electron micrographs revealed that the complex consists of two longitudinally aligned alpha/beta-tubulin heterodimers. The appearance of the complex was that of a kinked protofilament-like structure with a flat and a ribbed side. Deletion mapping of Op18 further demonstrated that (i) the function of the N-terminal part of the molecule is to 'cap' tubulin subunits to ensure the specificity of the complex and (ii) the complete C-terminal alpha-helical domain of Op18 is necessary and sufficient for stable Op18:tubulin complex formation. Together, our results suggest that besides sequestering tubulin, the structural features of Op18 enable the protein specifically to recognize microtubule ends to trigger catastrophes.
Figures
Similar articles
-
Dissociation of the tubulin-sequestering and microtubule catastrophe-promoting activities of oncoprotein 18/stathmin.Mol Biol Cell. 1999 Jan;10(1):105-18. doi: 10.1091/mbc.10.1.105. Mol Biol Cell. 1999. PMID: 9880330 Free PMC article.
-
Model for stathmin/OP18 binding to tubulin.EMBO J. 2000 Jan 17;19(2):213-22. doi: 10.1093/emboj/19.2.213. EMBO J. 2000. PMID: 10637225 Free PMC article.
-
Phosphorylation disrupts the central helix in Op18/stathmin and suppresses binding to tubulin.EMBO Rep. 2001 Jun;2(6):505-10. doi: 10.1093/embo-reports/kve105. EMBO Rep. 2001. PMID: 11415983 Free PMC article.
-
The oncoprotein 18/stathmin family of microtubule destabilizers.Curr Opin Cell Biol. 2002 Feb;14(1):18-24. doi: 10.1016/s0955-0674(01)00289-7. Curr Opin Cell Biol. 2002. PMID: 11792540 Review.
-
[The stathmin-tubulin interaction and the regulation of the microtubule assembly].Pathol Biol (Paris). 2003 Feb;51(1):33-8. doi: 10.1016/s0369-8114(02)00324-3. Pathol Biol (Paris). 2003. PMID: 12628290 Review. French.
Cited by
-
Global regulation of the interphase microtubule system by abundantly expressed Op18/stathmin.Mol Biol Cell. 2008 Jul;19(7):2897-906. doi: 10.1091/mbc.e08-01-0058. Epub 2008 Apr 23. Mol Biol Cell. 2008. PMID: 18434595 Free PMC article.
-
The contribution of αβ-tubulin curvature to microtubule dynamics.J Cell Biol. 2014 Nov 10;207(3):323-34. doi: 10.1083/jcb.201407095. J Cell Biol. 2014. PMID: 25385183 Free PMC article. Review.
-
Structural rearrangements in tubulin following microtubule formation.EMBO Rep. 2005 Mar;6(3):227-32. doi: 10.1038/sj.embor.7400360. EMBO Rep. 2005. PMID: 15731766 Free PMC article.
-
Stathmin and interfacial microtubule inhibitors recognize a naturally curved conformation of tubulin dimers.J Biol Chem. 2010 Oct 8;285(41):31672-81. doi: 10.1074/jbc.M110.141929. Epub 2010 Jul 30. J Biol Chem. 2010. PMID: 20675373 Free PMC article.
-
Deciphering the cellular functions of the Op18/Stathmin family of microtubule-regulators by plasma membrane-targeted localization.Mol Biol Cell. 2003 Sep;14(9):3716-29. doi: 10.1091/mbc.e03-03-0126. Epub 2003 May 29. Mol Biol Cell. 2003. PMID: 12972559 Free PMC article.
References
-
- Belmont L. and Mitchison, T.J. (1996a) Identification of a protein that interacts with tubulin dimers and increases the catastrophe rate of microtubules. Cell, 84, 623–631. - PubMed
-
- Belmont L. and Mitchison, T. (1996b) Catastrophic revelations about Op18/stathmin. Trends Biochem. Sci., 21, 197–198. - PubMed
-
- Bodenhausen G. and Ruben, D.J. (1980) Natural abundance nitrogen-15 NMR by enhanced heteronuclear spectroscopy. Chem. Phys. Lett., 69, 185–189.
-
- Bremer A. and Aebi,U. (1994) Negative staining. In Celis, J.E. (ed.), Cell Biology: A Laboratory Handbook. Academic Press Inc., London, UK, Vol. 2, pp. 126–133.
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
