

NF-kappaB determines localization and features of cell death in epidermis
- PMID: 10675350
- PMCID: PMC377441
- DOI: 10.1172/JCI7630
NF-kappaB determines localization and features of cell death in epidermis
Abstract
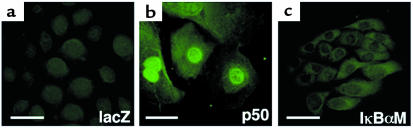
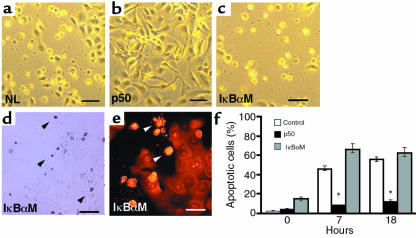
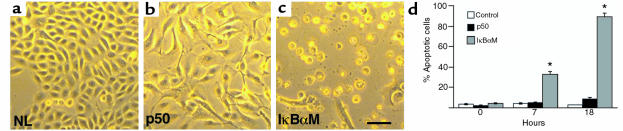
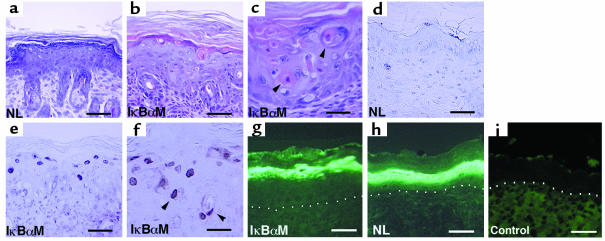
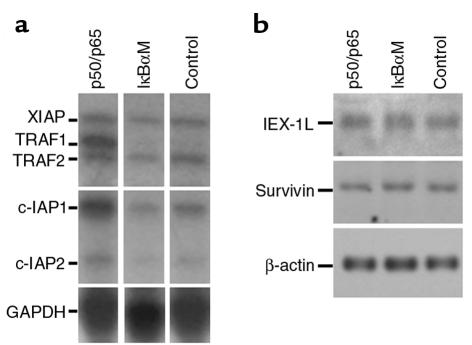
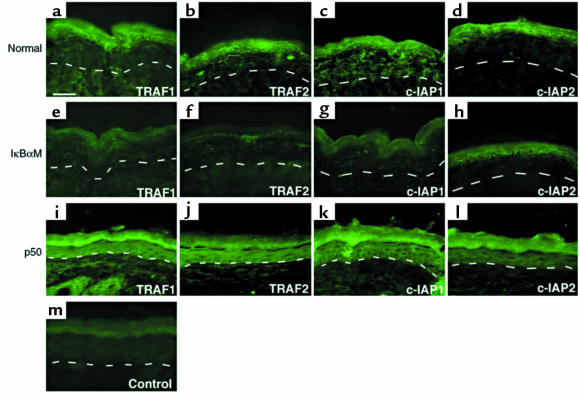
Specialized forms of physiologic cell death lacking certain characteristic morphologic features of apoptosis occur in terminally differentiating tissues, such as in the outer cell layers of epidermis. In these cell layers, NF-kappaB translocates from the cytoplasm to the nucleus and induces target gene expression. In light of its potent role in regulating apoptotic cell death in other tissues, NF-kappaB activation in these cells suggests that this transcription factor regulates cell death during terminal differentiation. Here, we show that NF-kappaB protects normal epithelial cells from apoptosis induced by both TNFalpha and Fas, whereas NF-kappaB blockade enhances susceptibility to death via both pathways. Expression of IkappaBalphaM under control of keratin promoter in transgenic mice caused a blockade of NF-kappaB function in the epidermis and provoked premature spontaneous cell death with apoptotic features. In normal tissue, expression of the known NF-kappaB-regulated antiapoptotic factors, TRAF1, TRAF2, c-IAP1, and c-IAP2, is most pronounced in outer epidermis. In transgenic mice, NF-kappaB blockade suppressed this expression, whereas NF-kappaB activation augmented it, consistent with regulation of cell death by these NF-kappaB effector proteins. These data identify a new role for NF-kappaB in preventing premature apoptosis in cells committed to undergoing physiologic cell death and indicate that, in stratified epithelium, such cell death normally proceeds via a distinct pathway that is resistant to NF-kappaB and its antiapoptotic target effector genes.
Figures
References
-
- Tamada Y, et al. Identification of programmed cell death in normal human skin tissues by using specific labelling of fragmented DNA. Br J Dermatol. 1994;131:521–524. - PubMed
-
- Polakowska R, Piacentini M, Bartlett R, Goldsmith L, Haake AR. Apoptosis in human skin development: morphogenesis, periderm, and stem cells. Dev Dyn. 1994;199:176–188. - PubMed
-
- Vaux DL, Korsmeyer SJ. Cell death in development. Cell. 1999;96:245–254. - PubMed
-
- Haake AR, Polakowska RR. Cell death by apoptosis in epidermal biology. J Invest Dermatol. 1993;101:107–112. - PubMed
-
- Hill LL, et al. Fas ligand: a sensor for DNA damage critical in skin cancer etiology. Science. 1999;285:898–900. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
