

Induction of topoisomerase I cleavage complexes by 1-beta -D-arabinofuranosylcytosine (ara-C) in vitro and in ara-C-treated cells
- PMID: 10677551
- PMCID: PMC26531
- DOI: 10.1073/pnas.97.4.1885
Induction of topoisomerase I cleavage complexes by 1-beta -D-arabinofuranosylcytosine (ara-C) in vitro and in ara-C-treated cells
Abstract
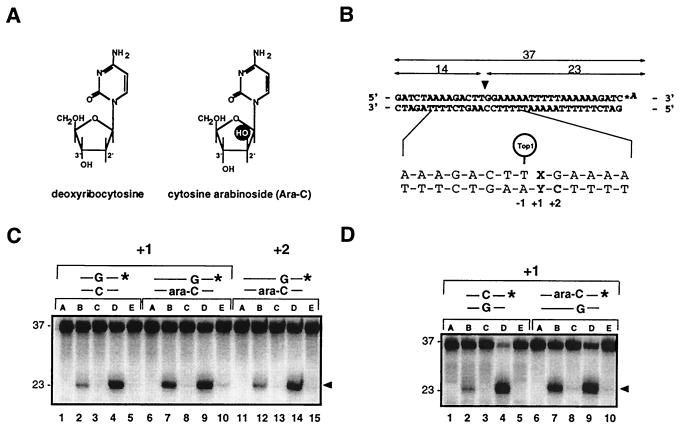
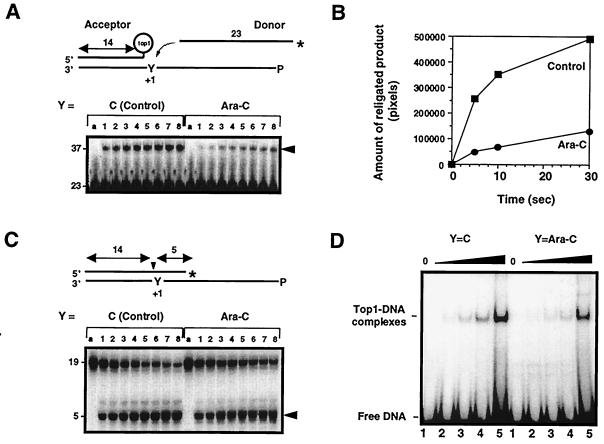
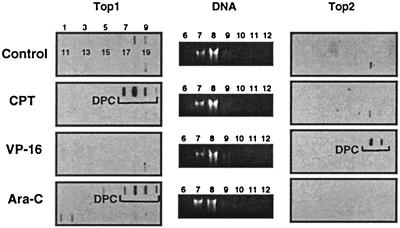
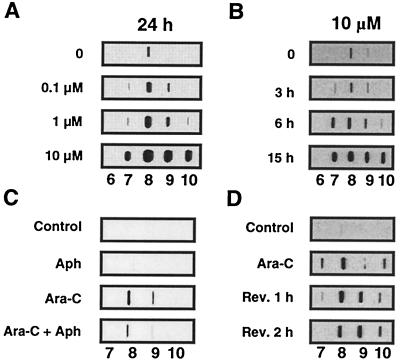
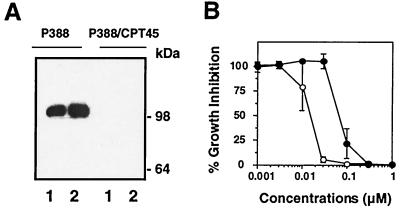
1-beta-d-Arabinofuranosylcytosine (Ara-C) is a nucleoside analog commonly used in the treatment of leukemias. Ara-C inhibits DNA polymerases and can be incorporated into DNA. Its mechanism of cytotoxicity is not fully understood. Using oligonucleotides and purified human topoisomerase I (top1), we found a 4- to 6-fold enhancement of top1 cleavage complexes when ara-C was incorporated at the +1 position (immediately 3') relative to a unique top1 cleavage site. This enhancement was primarily due to a reversible inhibition of top1-mediated DNA religation. Because ara-C incorporation is known to alter base stacking and sugar puckering at the misincorporation site and at the neighboring base pairs, the observed inhibition of religation at the ara-C site suggests the importance of the alignment of the 5'-hydroxyl end for religation with the phosphate group of the top1 phosphotyrosine bond. This study also demonstrates that ara-C treatment and DNA incorporation trap top1 cleavage complexes in human leukemia cells. Finally, we report that camptothecin-resistant mouse P388/CPT45 cells with no detectable top1 are crossresistant to ara-C, which suggests that top1 poisoning is a potential mechanism for ara-C cytotoxicity.
Figures
Similar articles
-
Structural impact of the leukemia drug 1-beta-D-arabinofuranosylcytosine (Ara-C) on the covalent human topoisomerase I-DNA complex.J Biol Chem. 2003 Apr 4;278(14):12461-6. doi: 10.1074/jbc.M212930200. Epub 2003 Jan 17. J Biol Chem. 2003. PMID: 12533542
-
Gemcitabine (2',2'-difluoro-2'-deoxycytidine), an antimetabolite that poisons topoisomerase I.Clin Cancer Res. 2002 Aug;8(8):2499-504. Clin Cancer Res. 2002. PMID: 12171875
-
Induction of topoisomerase I cleavage complexes by the vinyl chloride adduct 1,N6-ethenoadenine.J Biol Chem. 1998 Oct 16;273(42):27245-9. doi: 10.1074/jbc.273.42.27245. J Biol Chem. 1998. PMID: 9765247
-
Structural basis for topoisomerase I inhibition by nucleoside analogs.Nucleosides Nucleotides Nucleic Acids. 2003 May-Aug;22(5-8):653-8. doi: 10.1081/NCN-120022604. Nucleosides Nucleotides Nucleic Acids. 2003. PMID: 14565246 Review.
-
Diversity of DNA topoisomerases I and inhibitors.Biochimie. 1998 Mar;80(3):255-70. doi: 10.1016/s0300-9084(98)80008-4. Biochimie. 1998. PMID: 9615865 Review.
Cited by
-
TDP1 repairs nuclear and mitochondrial DNA damage induced by chain-terminating anticancer and antiviral nucleoside analogs.Nucleic Acids Res. 2013 Sep;41(16):7793-803. doi: 10.1093/nar/gkt483. Epub 2013 Jun 17. Nucleic Acids Res. 2013. PMID: 23775789 Free PMC article.
-
Entrapment of DNA topoisomerase-DNA complexes by nucleotide/nucleoside analogs.Cancer Drug Resist. 2019;2(4):994-1001. doi: 10.20517/cdr.2019.95. Epub 2019 Dec 19. Cancer Drug Resist. 2019. PMID: 31930190 Free PMC article.
-
DNA and RNA Cleavage Complexes and Repair Pathway for TOP3B RNA- and DNA-Protein Crosslinks.Cell Rep. 2020 Dec 29;33(13):108569. doi: 10.1016/j.celrep.2020.108569. Cell Rep. 2020. PMID: 33378676 Free PMC article.
-
Increased Expression of Micro-RNA-23a Mediates Chemoresistance to Cytarabine in Acute Myeloid Leukemia.Cancers (Basel). 2020 Feb 20;12(2):496. doi: 10.3390/cancers12020496. Cancers (Basel). 2020. PMID: 32093419 Free PMC article.
-
Oligonucleotides Carrying Nucleoside Antimetabolites as Potential Prodrugs.Curr Med Chem. 2023;30(11):1304-1319. doi: 10.2174/0929867328666211129124039. Curr Med Chem. 2023. PMID: 34844535 Free PMC article. Review.
References
-
- Hsieh T, Lee M P, Brown S D. Adv Pharmacol. 1994;29:191–200. - PubMed
-
- Wang J C. Annu Rev Biochem. 1996;65:635–692. - PubMed
-
- Champoux J. In: Mechanistic Aspects of Type-I Topoisomerases. Wang J C, Cozarelli N R, editors. Plainview, NY: Cold Spring Harbor Lab. Press; 1990. pp. 217–242.
-
- Gupta M, Fujimori A, Pommier Y. Biochim Biophys Acta. 1995;1262:1–14. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
