

Borrelia burgdorferi gene expression in vivo and spirochete pathogenicity
- PMID: 10678930
- PMCID: PMC97271
- DOI: 10.1128/IAI.68.3.1222-1230.2000
Borrelia burgdorferi gene expression in vivo and spirochete pathogenicity
Abstract
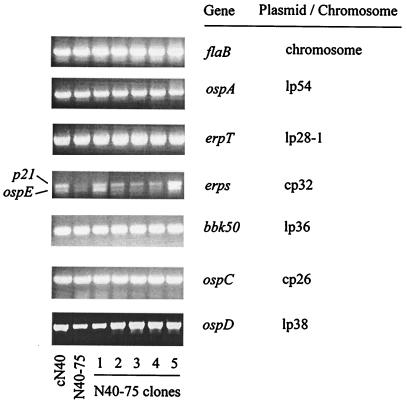
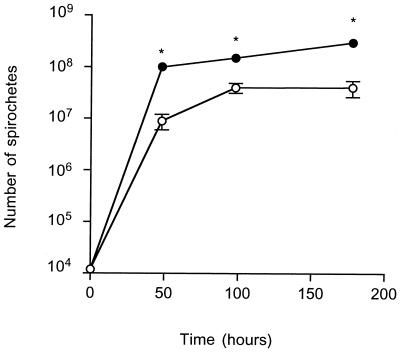
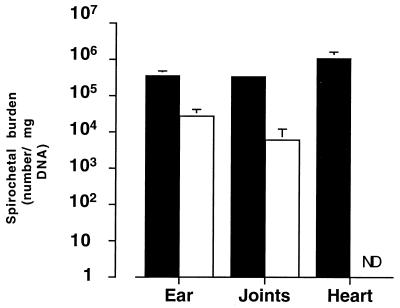
Borrelia burgdorferi spirochetes that do not cause arthritis or carditis were developed and used to investigate Lyme disease pathogenesis. A clonal isolate of B. burgdorferi N40 (cN40), which induces disease in C3H/HeN (C3H) mice, was repeatedly passaged in vitro to generate nonpathogenic spirochetes. The passage 75 isolate (N40-75) was infectious for C3H mice but did not cause arthritis or carditis, and spirochetes were at low levels or absent in the joints or hearts, respectively. N40-75 could, however, cause disease in severe combined immunodeficient (SCID) mice, suggesting that the response in immunocompetent mice prevented effective spirochete dissemination and the subsequent development of arthritis and carditis. Administration of immune sera at 4 days after spirochete challenge aborted N40-75, but not cN40, infection in SCID mice. A B. burgdorferi genomic expression library was differentially probed with sera from cN40- and N40-75-infected mice, to identify genes that may not be effectively expressed by N40-75 in vivo. N40-75 was defective in the up-regulation of several genes that are preferentially expressed during mammalian infection, including dbpAB, bba64, and genes that map to the cp32 family of plasmids. These data suggest that adaptation and gene expression may be required for B. burgdorferi to effectively colonize the host, evade humoral responses, and cause disease.
Figures



References
-
- Akins D R, Porcella S F, Popova T G, Shevchenko D, Baker S I, Li M, Norgard M V, Radolf J D. Evidence for in vivo but not in vitro expression of a Borrelia burgdorferi outer surface protein F (OspF) homologue. Mol Microbiol. 1995;18:507–520. - PubMed
-
- Armstrong A L, Barthold S W, Persing D H, Beck D S. Carditis in Lyme disease susceptible and resistant strains of laboratory mice infected with Borrelia burgdorferi. Am J Trop Med Hyg. 1992;47:249–258. - PubMed
-
- Barthold S W, Beck D S, Hansen G M, Terwilliger G A, Moody K D. Lyme borreliosis in selected strains and ages of laboratory mice. J Infect Dis. 1990;162:133–138. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
