

Influence of synthetic antiendotoxin peptides on lipopolysaccharide (LPS) recognition and LPS-induced proinflammatory cytokine responses by cells expressing membrane-bound CD14
- PMID: 10678985
- PMCID: PMC97326
- DOI: 10.1128/IAI.68.3.1655-1663.2000
Influence of synthetic antiendotoxin peptides on lipopolysaccharide (LPS) recognition and LPS-induced proinflammatory cytokine responses by cells expressing membrane-bound CD14
Abstract
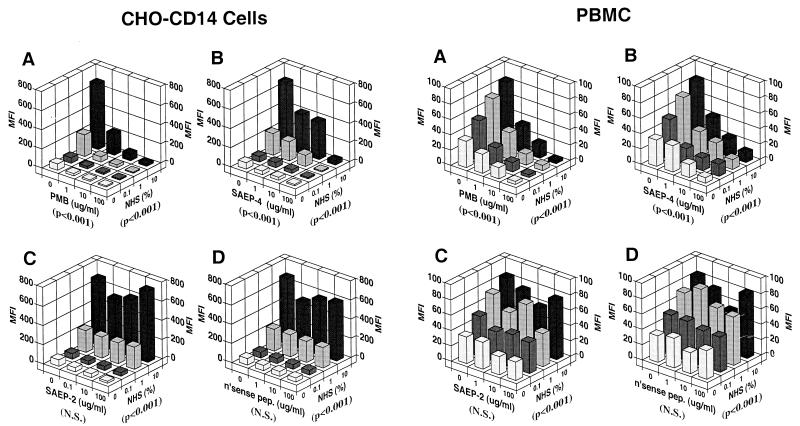
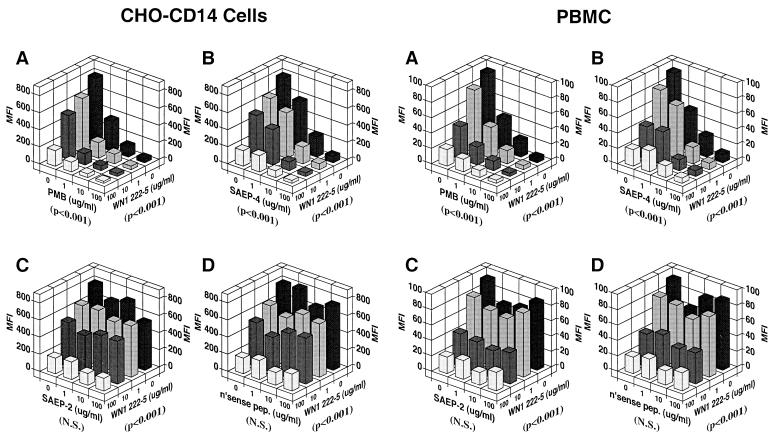
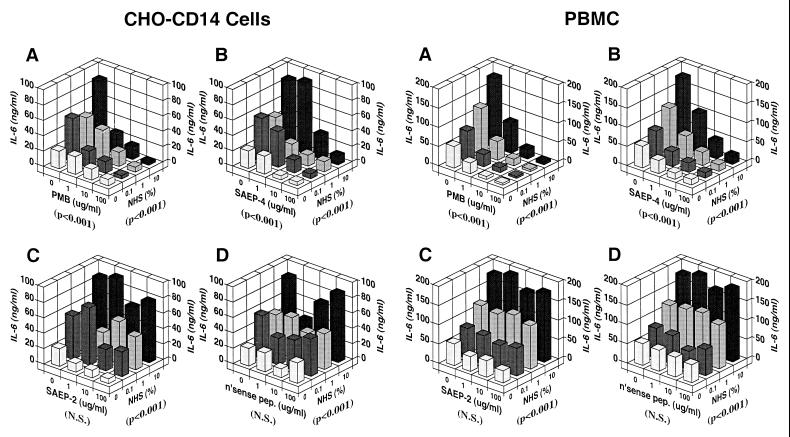
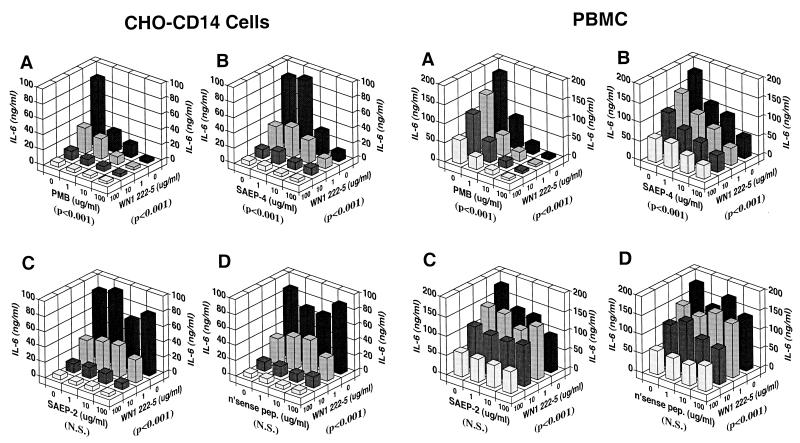
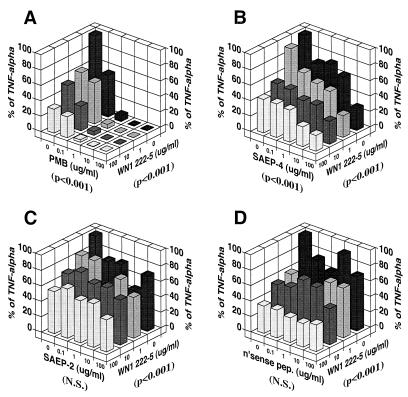
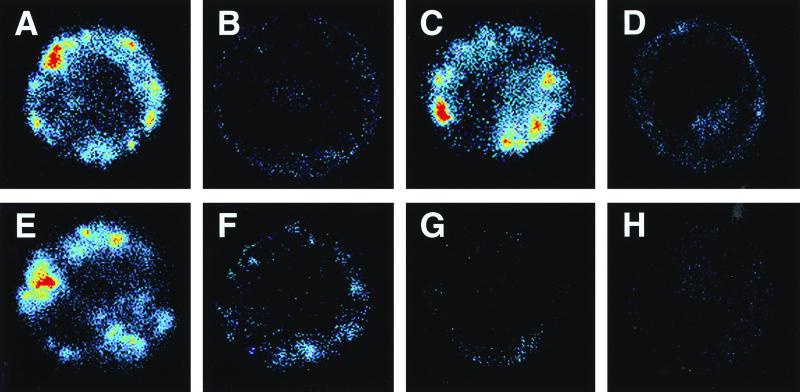
Lipopolysaccharides (LPS) are proinflammatory bacterial products implicated in the pathogenesis of gram-negative sepsis and septic shock. Polymyxin B (PMB), a cyclic, cationic peptide antibiotic, inhibits biological activities of LPS through high-affinity binding to the lipid A moiety. Small synthetic peptides have been designed to mimic the primary and secondary structures of PMB to determine structural requirements for binding and detoxification of lipid A and to assess possible therapeutic potential. The purpose of this study was to compare and contrast the endotoxin-neutralizing activities of two synthetic antiendotoxin peptides (SAEP-2 and SAEP-4), PMB, and an LPS core-specific monoclonal antibody (MAb), WN1 222-5, based on their abilities to inhibit CD14-mediated target cell uptake of fluorescein isothiocyanate (FITC)-conjugated LPS, detected by flow cytometry and confocal microscopy, and LPS-induced production of the proinflammatory cytokines, interleukin-6 (IL-6) and tumor necrosis factor alpha (TNF-alpha), as measured by bioassays. PMB and SAEP-4 produced dose-dependent inhibition of FITC-LPS uptake by CD14-transfected Chinese hamster ovary fibroblasts (CHO-CD14 cells) and by human peripheral blood mononuclear cells. The anti-LPS MAb, WN1 222-5, also blocked LPS uptake by these cells and synergized with PMB and SAEP-4. LPS-induced IL-6 release was inhibited by PMB, SAEP-4, and MAb WN1 222-5, and these inhibitory activities were additive or synergistic. LPS-induced TNF-alpha release by PBMC was also inhibited by PMB and SAEP-4 alone and in combination with anti-LPS MAb. SAEP-2, in contrast, produced comparatively minor decrements in cellular uptake of LPS and LPS-induced cytokine responses, and did so only in the absence of serum, while a nonsense peptide exerted no discernible inhibitory effect on LPS uptake or LPS-induced cytokine expression in the presence or absence of serum. Thus, PMB and SAEP-4, like the LPS-reactive MAb, WN1 222-5, block proinflammatory activities of LPS in part by preventing LPS recognition by membrane-bound CD14-expressing target cells. Differences in peptide structure, however, like those exemplified by SAEP-2 and SAEP-4, may differentially affect the endotoxin-neutralizing potency of these peptides despite similar binding activity against lipid A, reflecting possible differences in peptide solubility or peptide regulation of intracellular signal transduction.
Figures

Similar articles
-
Involvement of lipopolysaccharide binding protein, CD14, and Toll-like receptors in the initiation of innate immune responses by Treponema glycolipids.J Immunol. 2000 Sep 1;165(5):2683-93. doi: 10.4049/jimmunol.165.5.2683. J Immunol. 2000. PMID: 10946299
-
Dual effects of LPS antibodies on cellular uptake of LPS and LPS-induced proinflammatory functions.J Immunol. 1997 Oct 1;159(7):3519-30. J Immunol. 1997. PMID: 9317151
-
A synthetic peptide derived from bactericidal/permeability-increasing protein neutralizes endotoxin in vitro and in vivo.Int Immunopharmacol. 2004 Apr;4(4):527-37. doi: 10.1016/j.intimp.2004.02.004. Int Immunopharmacol. 2004. PMID: 15099530
-
Inhibition of lipopolysaccharide-induced TNF-alpha production by semisynthetic polymyxin-B conjugated dextran.Biotechnol Ther. 1994-1995;5(3-4):137-62. Biotechnol Ther. 1994. PMID: 8608324 Review.
-
Antiendotoxin strategies for the prevention and treatment of septic shock. New approaches and future directions.Drugs. 1998 Apr;55(4):497-508. doi: 10.2165/00003495-199855040-00002. Drugs. 1998. PMID: 9561340 Review.
Cited by
-
Nona-D-arginine amide suppresses corneal cytokines in Pseudomonas aeruginosa keratitis.Cornea. 2010 Nov;29(11):1308-14. doi: 10.1097/ICO.0b013e3181ca3a69. Cornea. 2010. PMID: 20802321 Free PMC article.
-
Schistosome larvae stimulate macrophage cytokine production through TLR4-dependent and -independent pathways.Int Immunol. 2005 Nov;17(11):1409-18. doi: 10.1093/intimm/dxh319. Epub 2005 Sep 26. Int Immunol. 2005. PMID: 16186163 Free PMC article.
-
Receptors, mediators, and mechanisms involved in bacterial sepsis and septic shock.Clin Microbiol Rev. 2003 Jul;16(3):379-414. doi: 10.1128/CMR.16.3.379-414.2003. Clin Microbiol Rev. 2003. PMID: 12857774 Free PMC article. Review.
-
Phylogeny and expression of ADAM10 and ADAM17 homologs in lamprey.Fish Physiol Biochem. 2023 Apr;49(2):321-334. doi: 10.1007/s10695-023-01184-7. Epub 2023 Mar 25. Fish Physiol Biochem. 2023. PMID: 36964830
-
Cecropin B enhances betalactams activities in experimental rat models of gram-negative septic shock.Ann Surg. 2004 Feb;239(2):251-6. doi: 10.1097/01.sla.0000108673.25385.03. Ann Surg. 2004. PMID: 14745334 Free PMC article.
References
-
- Coyne C P, Fenwick B W. Inhibition of lipopolysaccharide-induced macrophage tumor necrosis factor-α synthesis by polymyxin B sulfate. Am J Vet Res. 1993;54:305–314. - PubMed
-
- Demitri M T, Velucchi M, Bracci L, Rustici A, Porro M, Villa P, Ghezzi P. Inhibition of LPS-induced systemic and local TNF production by a synthetic anti-endotoxin peptide (SAEP-2) J Endotoxin Res. 1996;3:445–454.
-
- Evans T J, Carpenter A, Moyes D, Martin R, Cohen J. Protective effects of a recombinant amino-terminal fragment of human bactericidal/permeability-increasing protein in an animal model of gram-negative sepsis. J Infect Dis. 1995;171:153–160. - PubMed
-
- Golenbock D T, Liu Y, Millham F H, Freeman M W, Zoeller R A. Surface expression of human CD14 in Chinese hamster ovary fibroblasts imparts macrophage-like responsiveness to bacterial endotoxin. J Biol Chem. 1993;268:22055–22059. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
