

Essential functions of protein tyrosine phosphatases PTP2 and PTP3 and RIM11 tyrosine phosphorylation in Saccharomyces cerevisiae meiosis and sporulation
- PMID: 10679022
- PMCID: PMC14801
- DOI: 10.1091/mbc.11.2.663
Essential functions of protein tyrosine phosphatases PTP2 and PTP3 and RIM11 tyrosine phosphorylation in Saccharomyces cerevisiae meiosis and sporulation
Abstract
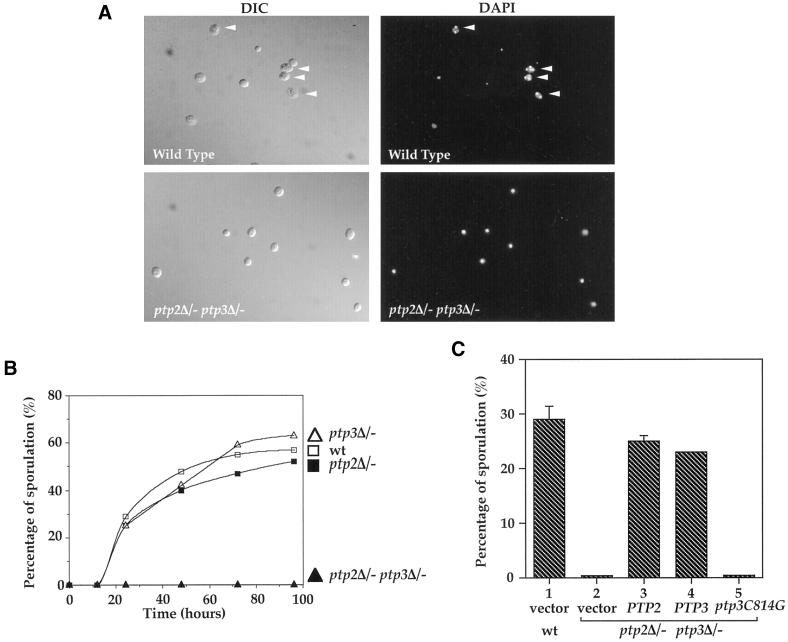
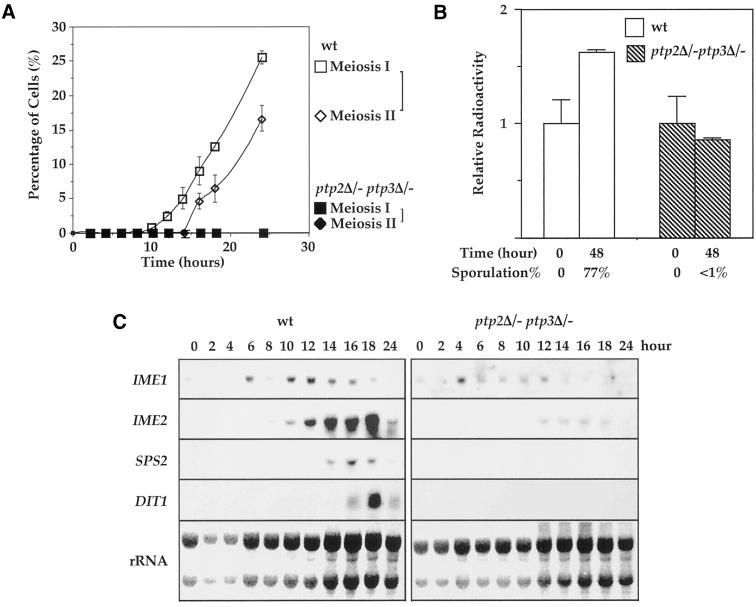
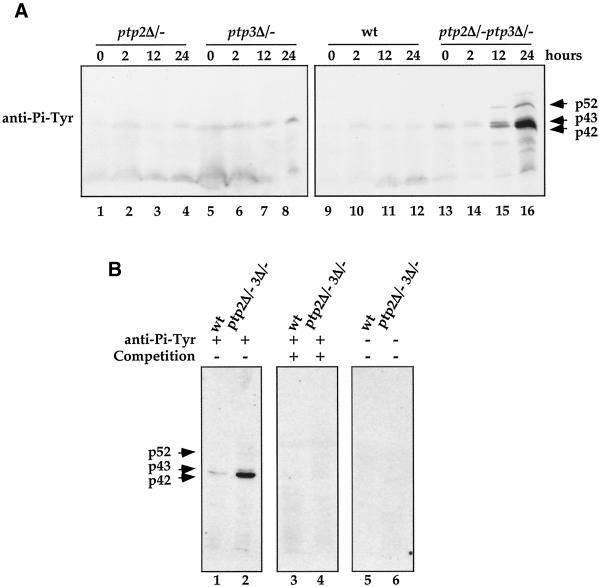
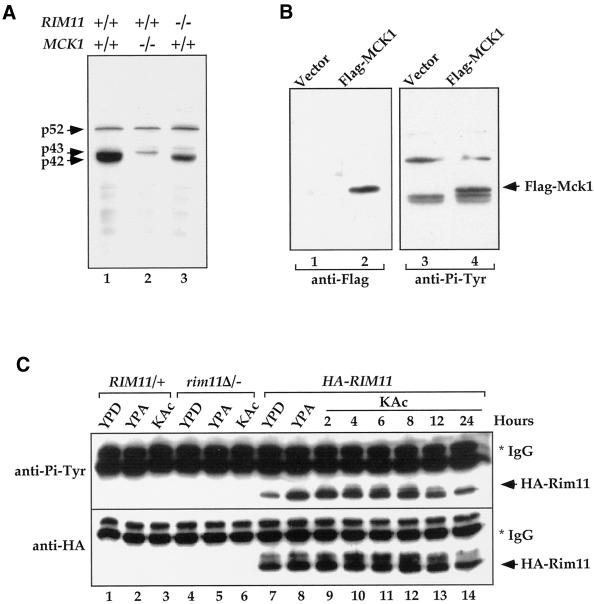
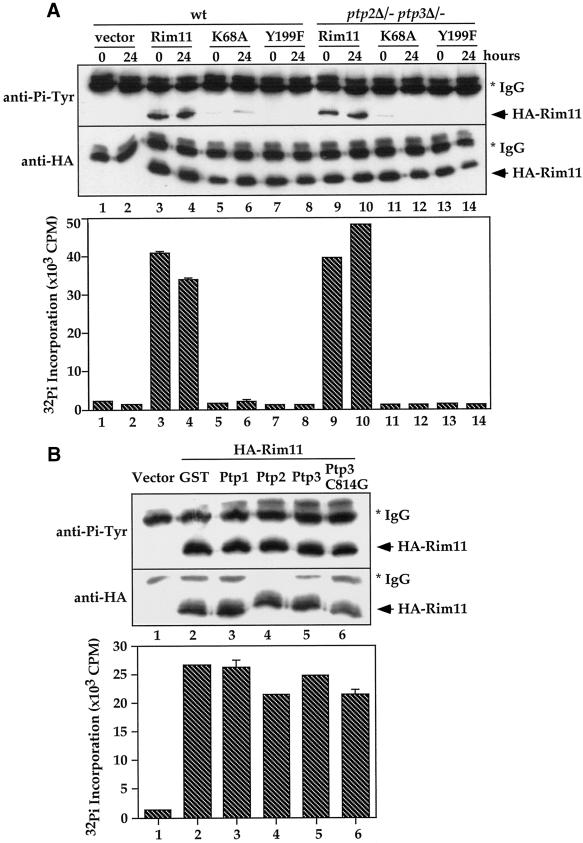
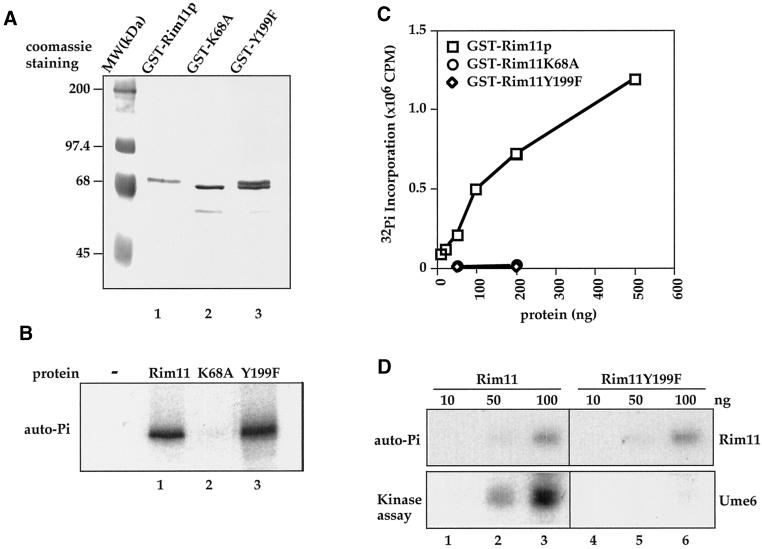
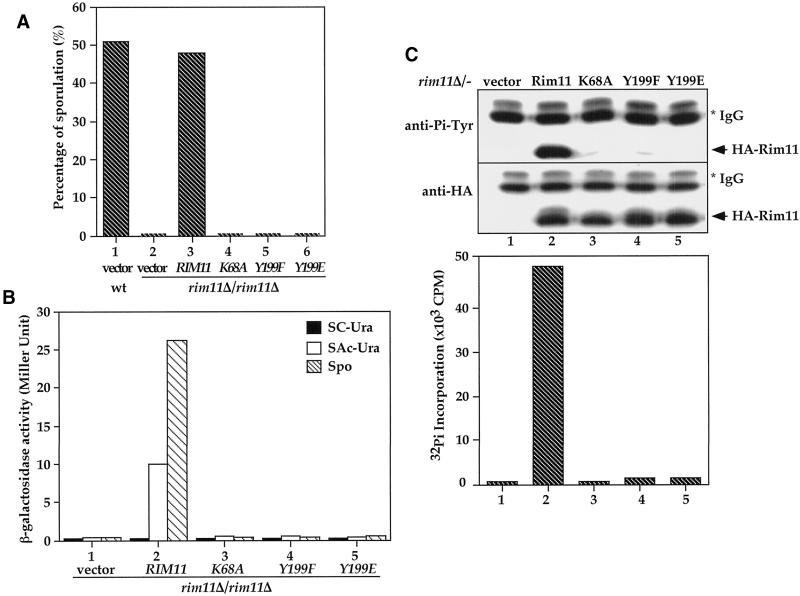
Tyrosine phosphorylation plays a central role in eukaryotic signal transduction. In yeast, MAP kinase pathways are regulated by tyrosine phosphorylation, and it has been speculated that other biochemical processes may also be regulated by tyrosine phosphorylation. Previous genetic and biochemical studies demonstrate that protein tyrosine phosphatases (PTPases) negatively regulate yeast MAP kinases. Here we report that deletion of PTP2 and PTP3 results in a sporulation defect, suggesting that tyrosine phosphorylation is involved in regulation of meiosis and sporulation. Deletion of PTP2 and PTP3 blocks cells at an early stage of sporulation before premeiotic DNA synthesis and induction of meiotic-specific genes. We observed that tyrosine phosphorylation of several proteins, including 52-, 43-, and 42-kDa proteins, was changed in ptp2Deltaptp3Delta homozygous deletion cells under sporulation conditions. The 42-kDa tyrosine-phosphorylated protein was identified as Mck1, which is a member of the GSK3 family of protein kinases and previously known to be phosphorylated on tyrosine. Mutation of MCK1 decreases sporulation efficiency, whereas mutation of RIM11, another GSK3 member, specifically abolishes sporulation; therefore, we investigated regulation of Rim11 by Tyr phosphorylation during sporulation. We demonstrated that Rim11 is phosphorylated on Tyr-199, and the Tyr phosphorylation is essential for its in vivo function, although Rim11 appears not to be directly regulated by Ptp2 and Ptp3. Biochemical characterizations indicate that tyrosine phosphorylation of Rim11 is essential for the activity of Rim11 to phosphorylate substrates. Our data demonstrate important roles of protein tyrosine phosphorylation in meiosis and sporulation
Figures
References
-
- Cobb MH, Goldsmith EJ. How MAP kinases are regulated. J Biol Chem. 1995;270:14843–14846. - PubMed
-
- Eisenmann DM, Kim SK. Signal transduction and cell fate specification during Caenorhabditis elegans vulval development. Curr Opin Genet Dev. 1994;4:508–516. - PubMed
-
- Friesen H, Lunz R, Doyle S, Segall J. Mutation of the SPS1-encoded protein kinase of Saccharomyces cerevisiae leads to defects in transcription and morphology during spore formation. Genes Dev. 1994;8:2162–2175. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
