

Electrostatic stress in catalysis: structure and mechanism of the enzyme orotidine monophosphate decarboxylase
- PMID: 10681441
- PMCID: PMC15746
- DOI: 10.1073/pnas.050417797
Electrostatic stress in catalysis: structure and mechanism of the enzyme orotidine monophosphate decarboxylase
Abstract
Orotidine 5'-monophosphate decarboxylase catalyzes the conversion of orotidine 5'-monophosphate to uridine 5'-monophosphate, the last step in biosynthesis of pyrimidine nucleotides. As part of a Structural Genomics Initiative, the crystal structures of the ligand-free and the6-azauridine 5'-monophosphate-complexed forms have been determined at 1.8 and 1.5 A, respectively. The protein assumes a TIM-barrel fold with one side of the barrel closed off and the other side binding the inhibitor. A unique array of alternating charges (Lys-Asp-Lys-Asp) in the active site prompted us to apply quantum mechanical and molecular dynamics calculations to analyze the relative contributions of ground state destabilization and transition state stabilization to catalysis. The remarkable catalytic power of orotidine 5'-monophosphate decarboxylase is almost exclusively achieved via destabilization of the reactive part of the substrate, which is compensated for by strong binding of the phosphate and ribose groups. The computational results are consistent with a catalytic mechanism that is characterized by Jencks's Circe effect.
Figures
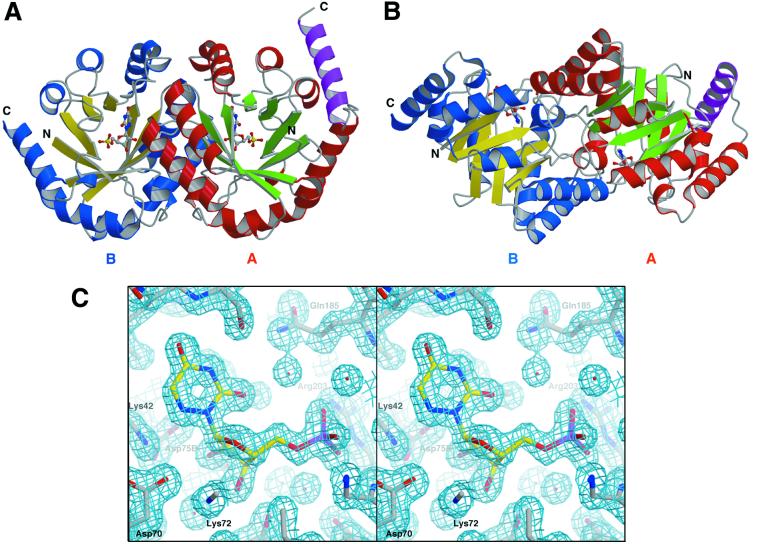
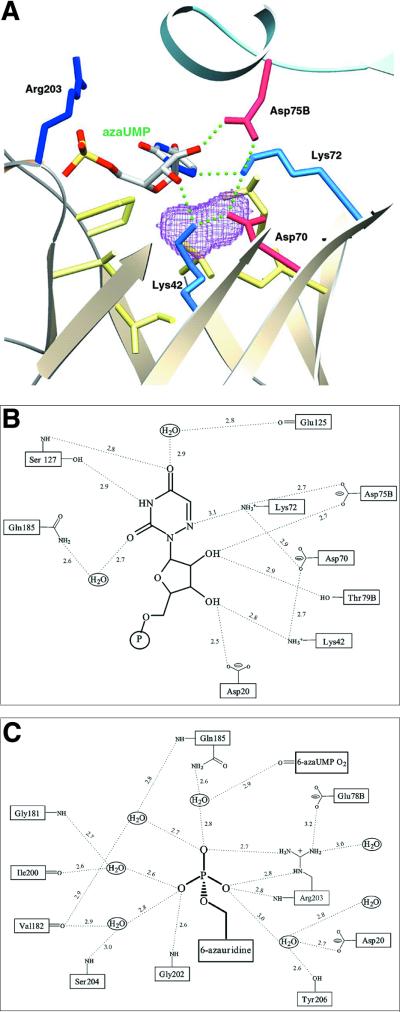
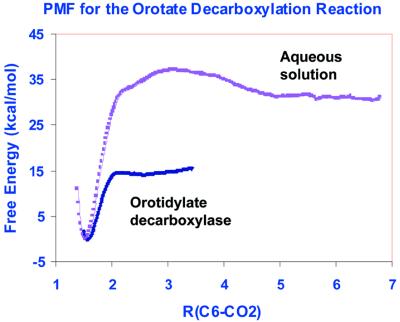
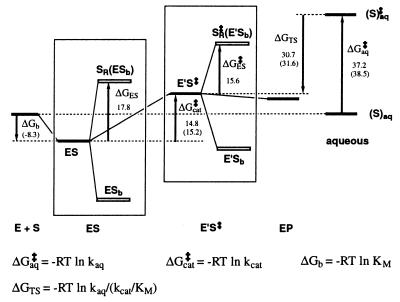
References
-
- Lieberman I, Kornberg A, Simms E S. J Biol Chem. 1955;215:403–415. - PubMed
-
- Radzicka A, Wolfenden R. Science. 1995;267:90–93. - PubMed
-
- Beak P, Siegel B. J Am Chem Soc. 1976;98:3601–3606. - PubMed
-
- Levine H L, Brody R S, Westheimer F H. Biochemistry. 1980;19:4993–4999. - PubMed
-
- Acheson S A, Bell J B, Jones M E, Wolfenden R. Biochemistry. 1990;29:3198–3202. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
