

Efficient gene transfer into human CD34(+) cells by a retargeted adenovirus vector
- PMID: 10684271
- PMCID: PMC111745
- DOI: 10.1128/jvi.74.6.2567-2583.2000
Efficient gene transfer into human CD34(+) cells by a retargeted adenovirus vector
Abstract
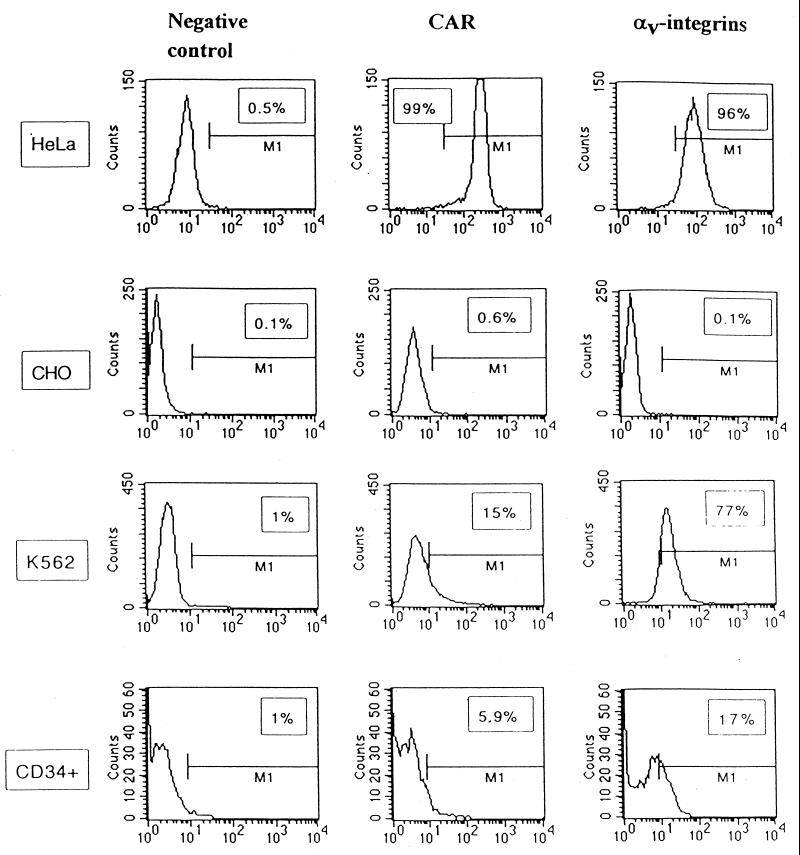
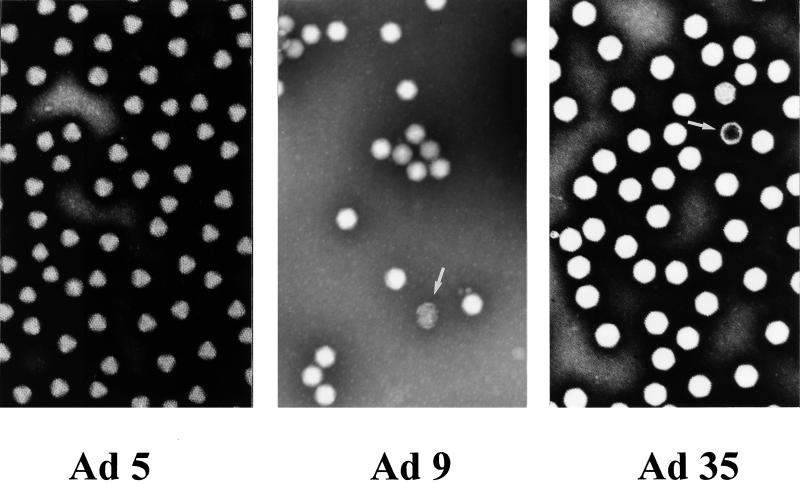
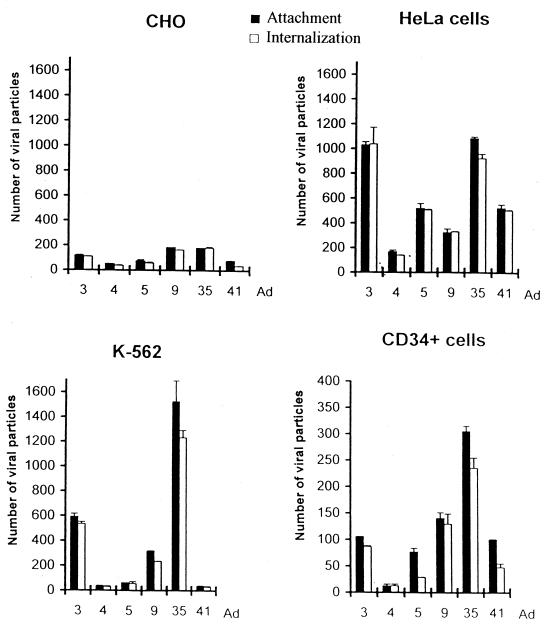
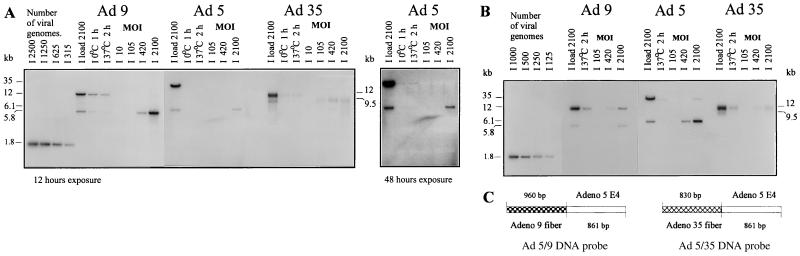
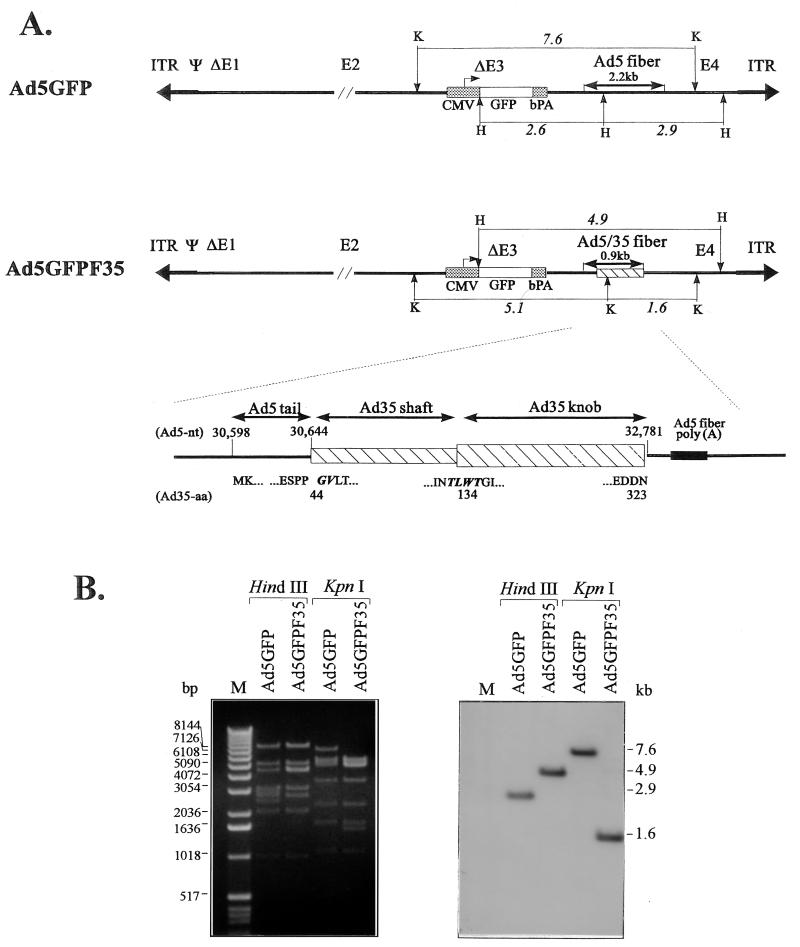
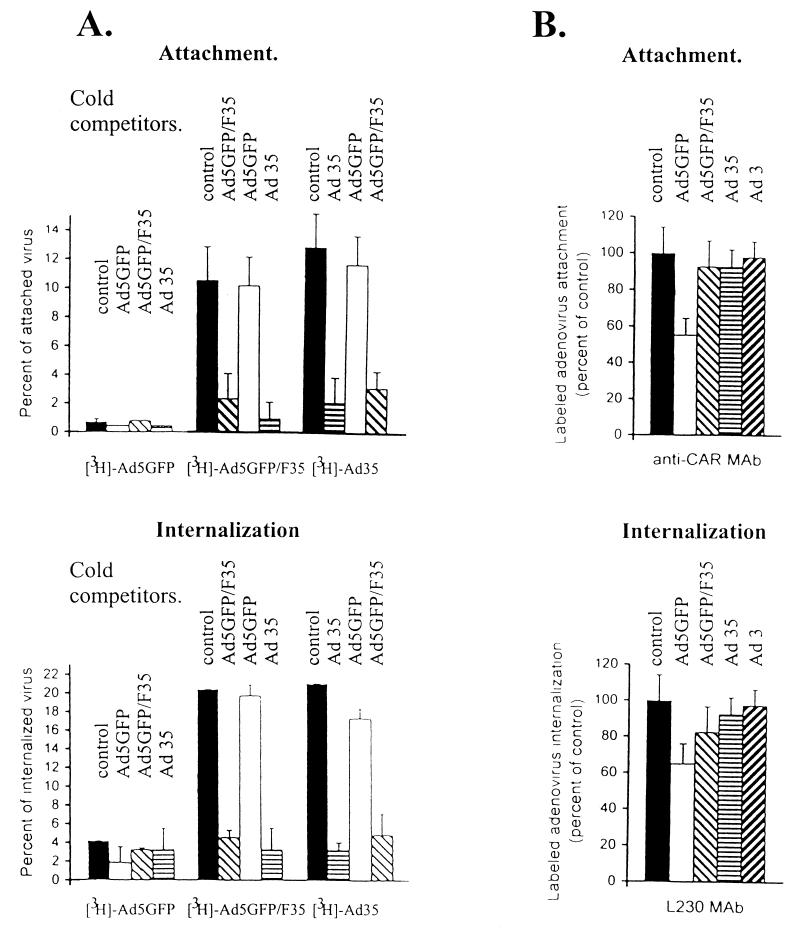
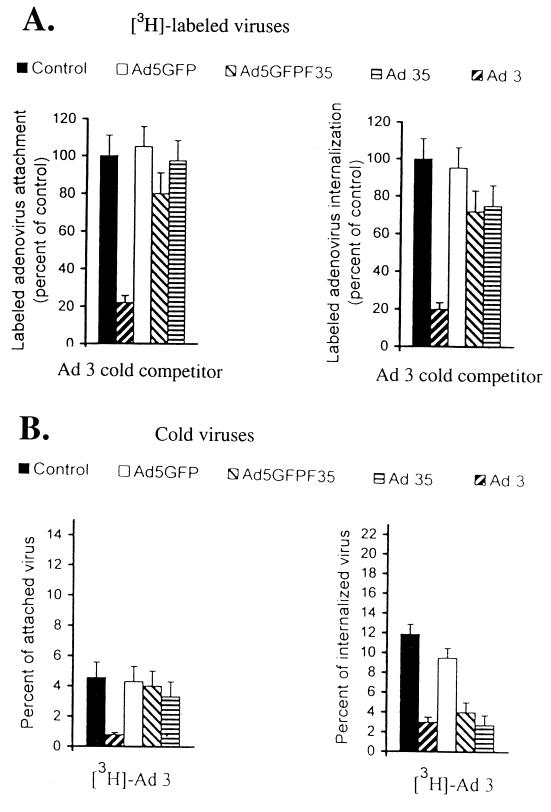
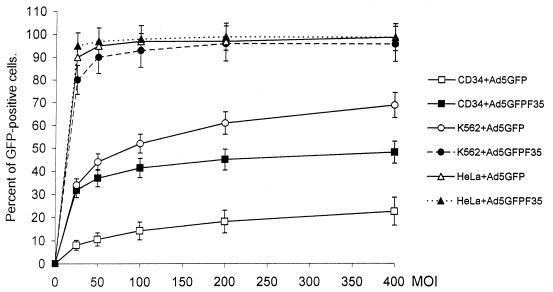
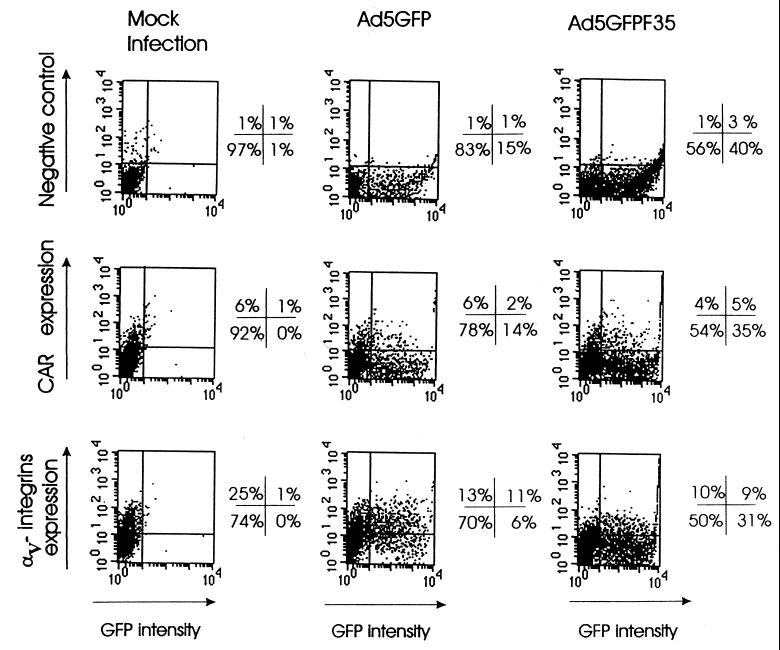
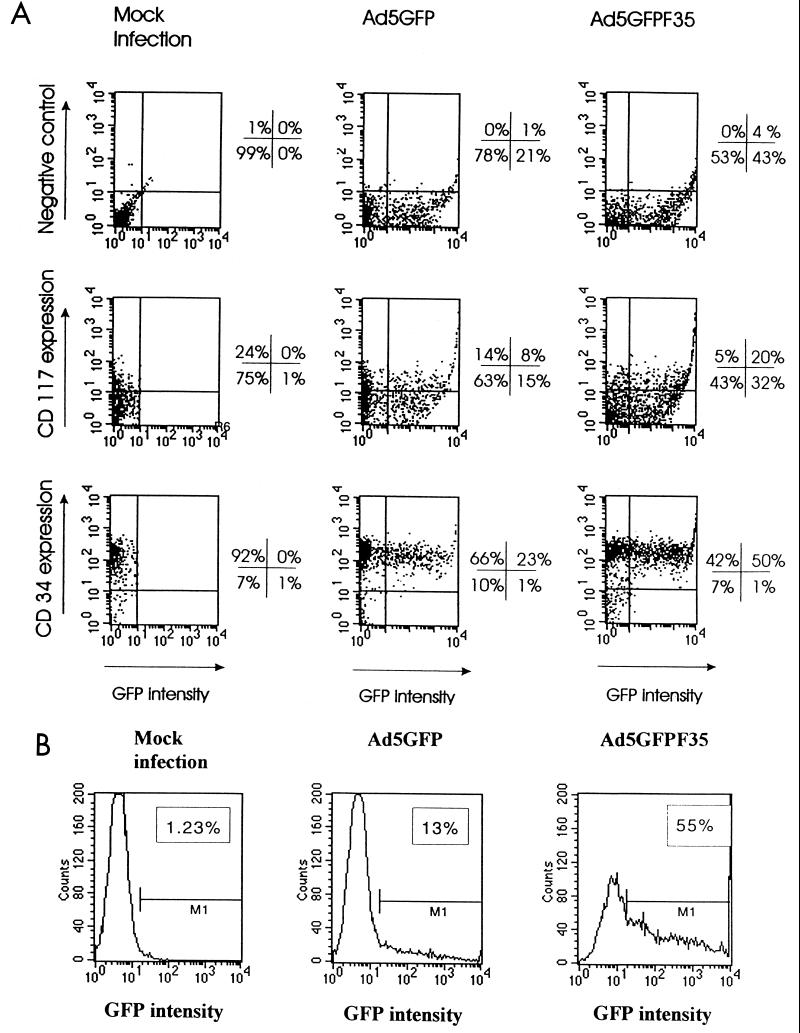
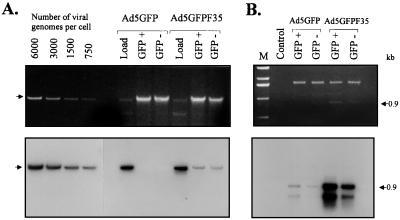
Efficient infection with adenovirus (Ad) vectors based on serotype 5 (Ad5) requires the presence of coxsackievirus-adenovirus receptors (CAR) and alpha(v) integrins on cells. The paucity of these cellular receptors is thought to be a limiting factor for Ad gene transfer into hematopoietic stem cells. In a systematic approach, we screened different Ad serotypes for interaction with noncycling human CD34(+) cells and K562 cells on the level of virus attachment, internalization, and replication. From these studies, serotype 35 emerged as the variant with the highest tropism for CD34(+) cells. A chimeric vector (Ad5GFP/F35) was generated which contained the short-shafted Ad35 fiber incorporated into an Ad5 capsid. This substitution was sufficient to transplant all infection properties from Ad35 to the chimeric vector. The retargeted, chimeric vector attached to a receptor different from CAR and entered cells by an alpha(v) integrin-independent pathway. In transduction studies, Ad5GFP/F35 expressed green fluorescent protein (GFP) in 54% of CD34(+) cells. In comparison, the standard Ad5GFP vector conferred GFP expression to only 25% of CD34(+) cells. Importantly, Ad5GFP transduction, but not Ad5GFP/F35, was restricted to a specific subset of CD34(+) cells expressing alpha(v) integrins. The actual transduction efficiency was even higher than 50% because Ad5GFP/F35 viral genomes were found in GFP-negative CD34(+) cell fractions, indicating that the cytomegalovirus promoter used for transgene expression was not active in all transduced cells. The chimeric vector allowed for gene transfer into a broader spectrum of CD34(+) cells, including subsets with potential stem cell capacity. Fifty-five percent of CD34(+) c-Kit(+) cells expressed GFP after infection with Ad5GFP/F35, whereas only 13% of CD34(+) c-Kit(+) cells were GFP positive after infection with Ad5GFP. These findings represent the basis for studies aimed toward stable gene transfer into hematopoietic stem cells.
Figures
References
-
- Bailey A, Mautner V. Phylogenetic relationships among adenovirus serotypes. Virology. 1994;205:438–452. - PubMed
-
- Becker P S, Nilsson S K, Li Z, Berrios V M, Dooner M S, Cooper C L, Hsieh C C, Quesenberry P J. Adhesion receptor expression by hematopoietic cell lines and murine progenitors: modulation by cytokines and cell cycle status. Exp Hematol. 1999;27:533–541. - PubMed
-
- Bergelson J M, Cunningham J A, Droguett G, Kurt-Jones E A, Krithivas A, Hong J S, Horwitz M S, Crowell R L, Finberg R W. Isolation of a common receptor for coxsackie B viruses and adenoviruses 2 and 5. Science. 1997;275:1320–1323. - PubMed
-
- Bodine D M, Moritz T, Donahue R E, Luskey B D, Kessler S W, Martin D I K, Orkin S H, Nienhuis A W, Williams D A. Long-term in vivo expansion of a murine adenosine deaminase gene in rhesus monkey hematopoietic cells of multiple lineages after retroviral mediated gene transfer into CD34+ bone marrow cells. Blood. 1993;82:1975–1980. - PubMed
-
- Bregni M, Shammah S, Malaffo F, Di Nicola M, Milanesi M. Adenovirus vectors for gene transduction into mobilized blood CD34+ cells. Gene Ther. 1998;5:465–472. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
