

Repression of the integrated papillomavirus E6/E7 promoter is required for growth suppression of cervical cancer cells
- PMID: 10684283
- PMCID: PMC111757
- DOI: 10.1128/jvi.74.6.2679-2686.2000
Repression of the integrated papillomavirus E6/E7 promoter is required for growth suppression of cervical cancer cells
Abstract
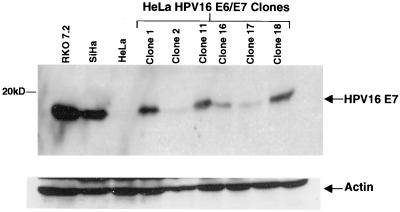
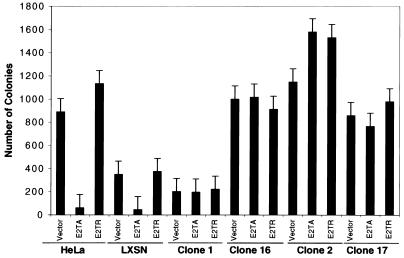
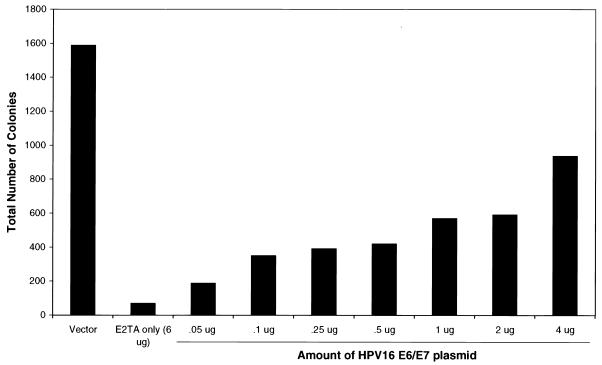
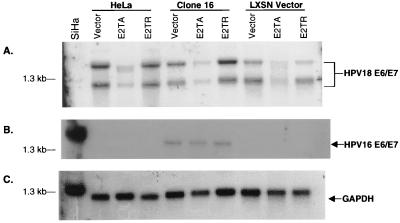
The human papillomavirus (HPV) E2 protein is an important regulator of viral E6 and E7 gene expression. E2 can repress the viral promoter for E6 and E7 expression as well as block progression of the cell cycle in cancer cells harboring the DNA of "high-risk" HPV types. Although the phenomenon of E2-mediated growth arrest of HeLa cells and other HPV-positive cancer cells has been well documented, the specific mechanism by which E2 affects cellular proliferation has not yet been elucidated. Here, we show that bovine papillomavirus (BPV) E2-induced growth arrest of HeLa cells requires the repression of the E6 and E7 promoter. This repression is specific for E2TA and not E2TR, a BPV E2 variant that lacks the N-terminal transactivation domain. We demonstrate that expression of HPV16 E6 and E7 from a heterologous promoter that is not regulated by E2 rescues HeLa cells from E2-mediated growth arrest. Our data indicate that the pathway of E2-mediated growth arrest of HeLa cells requires repression of E6 and E7 expression through an activity specified by the transactivation domain of E2TA.
Figures


References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
