

Promyelocytic leukemia protein (PML) and Daxx participate in a novel nuclear pathway for apoptosis
- PMID: 10684855
- PMCID: PMC2195846
- DOI: 10.1084/jem.191.4.631
Promyelocytic leukemia protein (PML) and Daxx participate in a novel nuclear pathway for apoptosis
Abstract
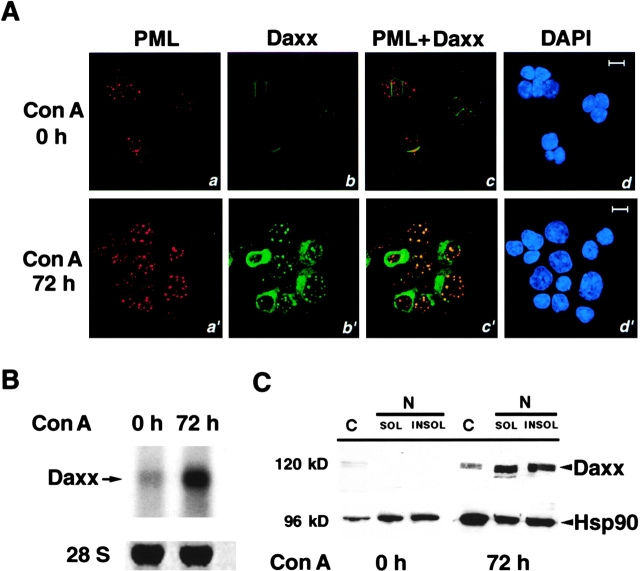
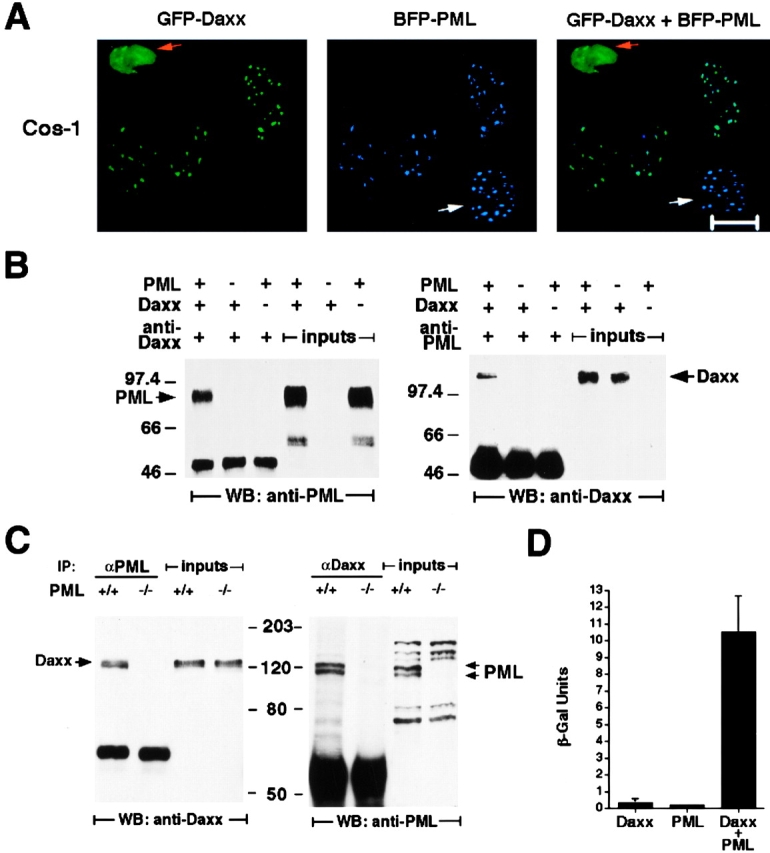
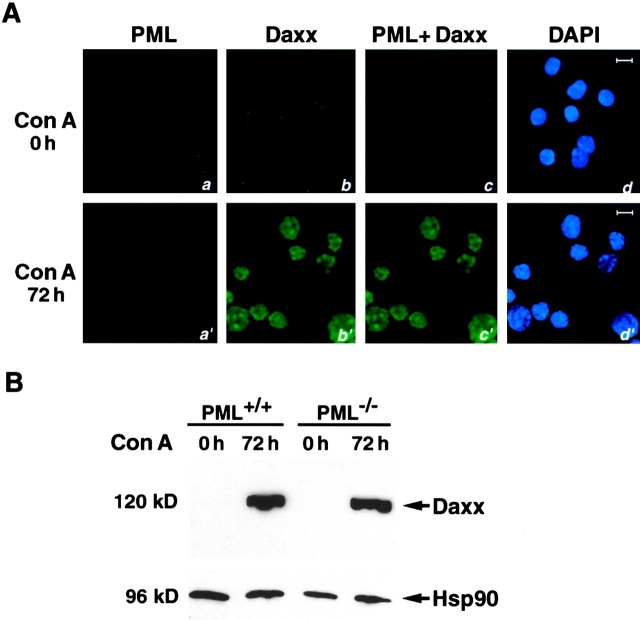
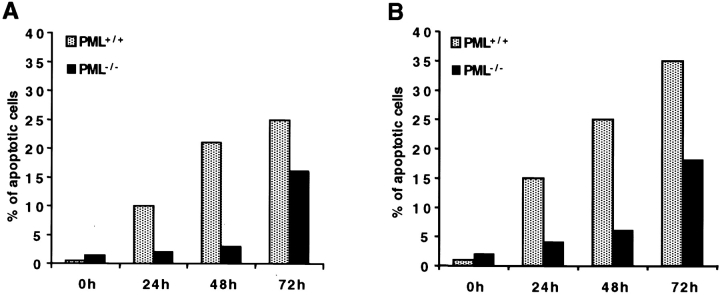
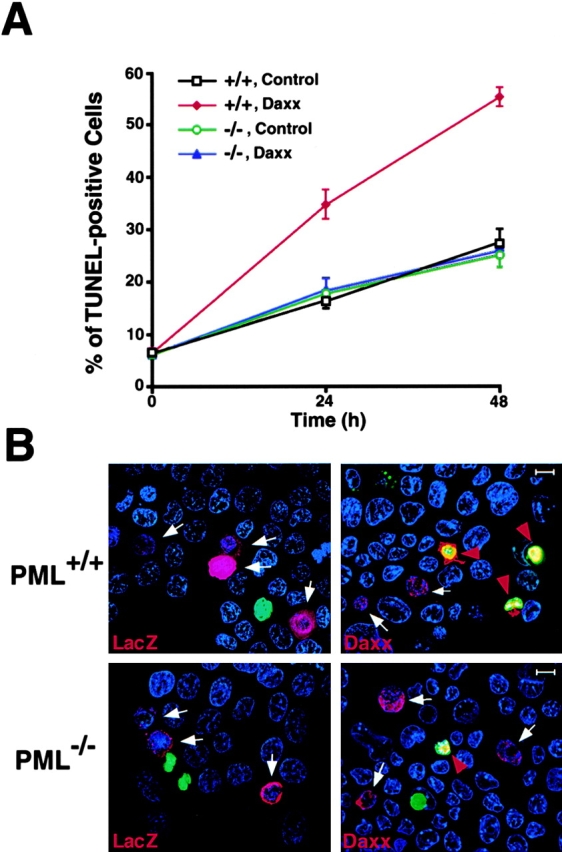
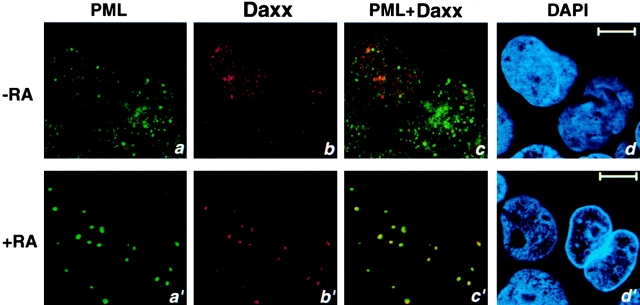
The promyelocytic leukemia protein (PML) gene of acute promyelocytic leukemia (APL) encodes a cell growth and tumor suppressor essential for multiple apoptotic signals. Daxx was identified as a molecule important for the cytoplasmic transduction of the Fas proapoptotic stimulus. Here, we show that upon mitogenic activation of mature splenic lymphocytes, Daxx is dramatically upregulated and accumulates in the PML nuclear body (NB) where PML and Daxx physically interact. In the absence of PML, Daxx acquires a dispersed nuclear pattern, and activation-induced cell death of splenocytes is profoundly impaired. PML inactivation results in the complete abrogation of the Daxx proapoptotic ability. In APL cells, Daxx is delocalized from the NB. Upon retinoic acid treatment, which induces disease remission in APL, Daxx relocalizes to the PML NBs. These results indicate that PML and Daxx cooperate in a novel NB-dependent pathway for apoptosis and shed new light in the role of PML in tumor suppression.
Figures

References
-
- Pandolfi P.P., Grignani F., Alcalay M., Mencarelli A., Biondi A., Lo Coco F., Pelicci P.G. Structure and origin of the acute promyelocytic leukemia myl/RARα cDNA and characterization of its retinoid-binding and transactivation properties. Oncogene. 1991;6:1285–1292. - PubMed
-
- de Thé H., Lavau C., Marchio A., Chomienne C., Degos L., Dejean A. The PML/RARα fusion mRNA generated by the t(15;17) translocation in acute promyelocytic leukemia encodes a functionally altered RAR. Cell. 1991;66:675–684. - PubMed
-
- Kakizuka A., Miller W.H., Jr., Umesono K., Warrell R.P., Jr., Frankel S.R., Murty V.V.V.S., Dmitrovsky E., Evans R.M. Chromosomal translocation t(15;17) in human acute promyelocytic leukemia fuses RARα with a novel putative transcription factor, PML. Cell. 1991;66:663–674. - PubMed
-
- Goddard A.D., Borrow P.S., Freemont P.S., Solomon E. Characterization of a zinc finger gene disrupted by the t(15;17) in acute promyelocytic leukemia. Science. 1991;254:1371–1374. - PubMed
-
- He L.Z., Merghoub T., Pandolfi P.P. In vivo analysis of the molecular pathogenesis of acute promyelocytic leukemia in the mouse and its therapeutic implications. Oncogene. 1999;18:5278–5292. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
