

Effects of RNA secondary structure on cellular antisense activity
- PMID: 10684928
- PMCID: PMC111043
- DOI: 10.1093/nar/28.6.1340
Effects of RNA secondary structure on cellular antisense activity
Abstract
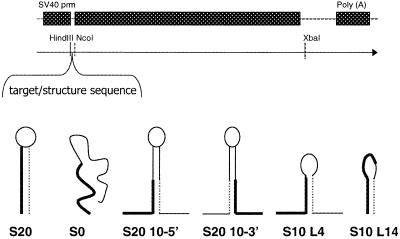
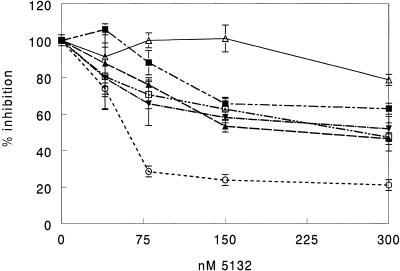
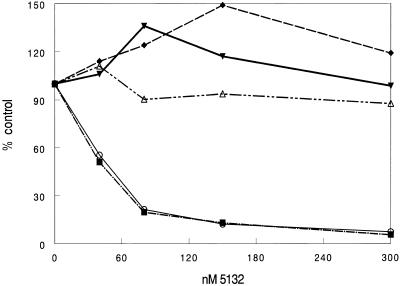
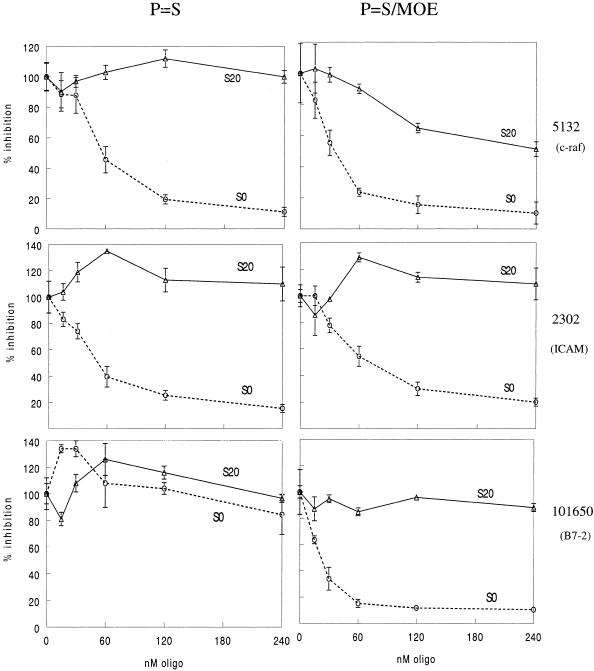
The secondary and tertiary structures of a mRNA are known to effect hybridization efficiency and potency of antisense oligonucleotides in vitro. Additional factors including oligonucleotide stability and cellular uptake are also thought to contribute to antisense potency in vivo. Each of these factors can be affected by the sequence of the oligonucleotide. Although mRNA structure is presumed to be a critical determinant of antisense activity in cells, to date little direct experimental evidence has addressed the significance of structure. In order to determine the importance of mRNA structure on antisense activity, oligonucleotide target sites were cloned into a luciferase reporter gene along with adjoining sequence to form known structures. This allowed us to study the effect of target secondary structure on oligonucleotide binding in the cellular environment without changing the sequence of the oligonucleotide. Our results show that structure does play a significant role in determining oligonucleotide efficacy in vivo. We also show that potency of oligonucleotides can be improved by altering chemistry to increase affinity for the mRNA target even in a region that is highly structured.
Figures
References
-
- Crooke S.T. (1998) Antisense Nucleic Acid Drug Dev., 8, 133–134. - PubMed
-
- Freier S.M. and Tinoco,I.J. (1975) Biochemistry, 14, 3310–3314. - PubMed
-
- Herschlag D. and Cech,T.R. (1990) Biochemistry, 29, 10159–10171. - PubMed
-
- Eckardt S., Romby,P. and Sczakiel,G. (1997) Biochemistry, 36, 12711–12721. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
