

Marking synaptic activity in dendritic spines with a calpain substrate exhibiting fluorescence resonance energy transfer
- PMID: 10688895
- PMCID: PMC15787
- DOI: 10.1073/pnas.040565597
Marking synaptic activity in dendritic spines with a calpain substrate exhibiting fluorescence resonance energy transfer
Abstract
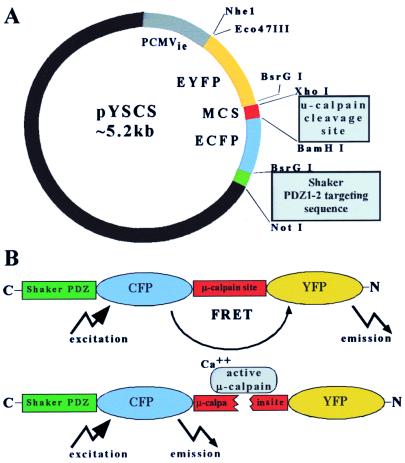
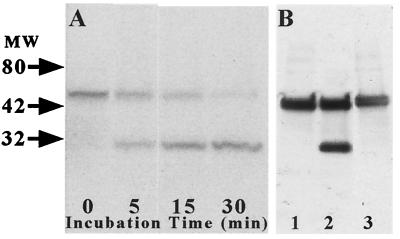
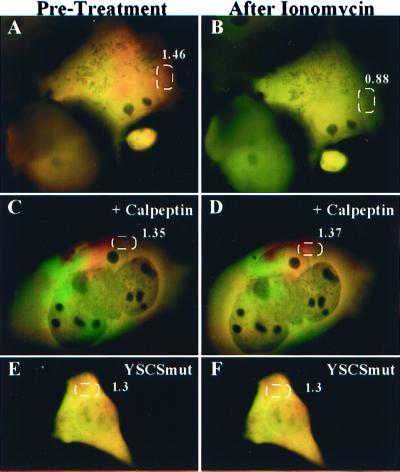
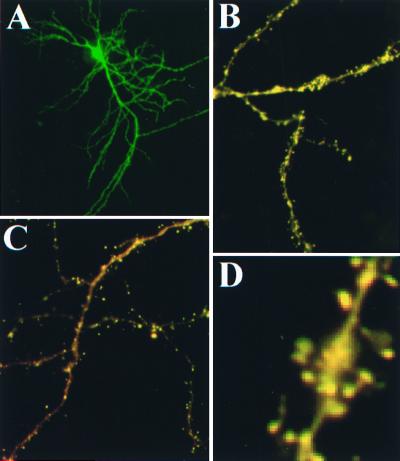
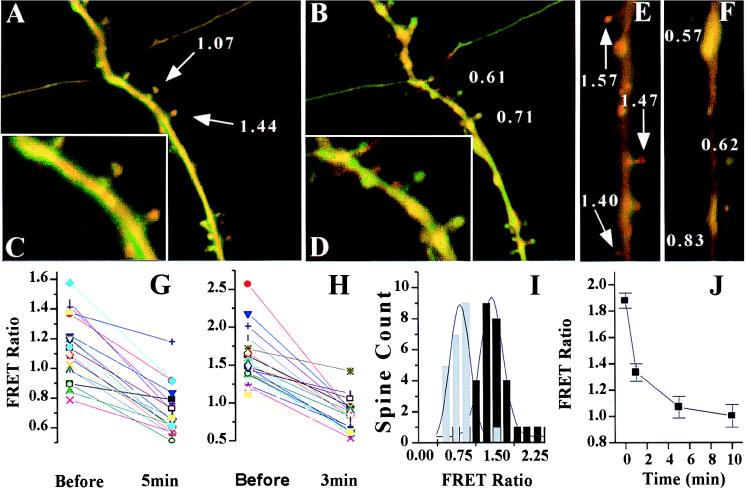
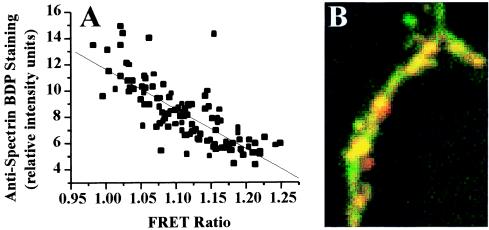
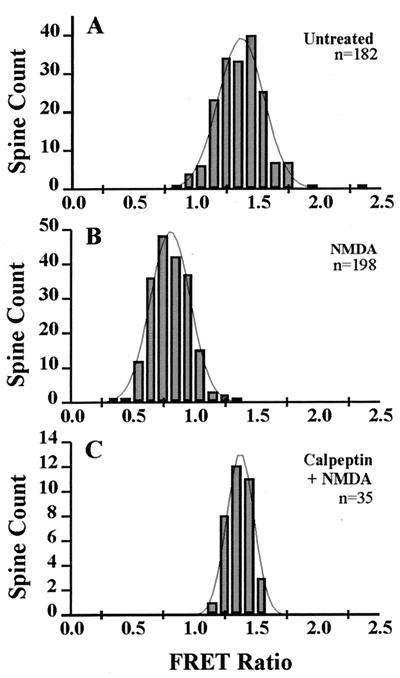
Excitatory synaptic activity can evoke transient and substantial elevations of postsynaptic calcium. Downstream effects of elevated calcium include the activation of the calcium-dependent protease calpain. We have developed a reagent that identifies dendritic spines in which calpain has been activated. A fusion protein was expressed that contained enhanced yellow and enhanced cyan fluorescent protein (EYFP and ECFP, respectively) linked by a peptide that included the micro-calpain cleavage site from alpha-spectrin. A PDZ-binding site fused to ECFP anchored this protein to postsynaptic densities. The fusion protein exhibited fluorescence resonance energy transfer (FRET), and diminution of FRET by proteolysis was used to localize calpain activity in situ by fluorescence microscopy. Incubation of the fusion protein with calpain in the presence of calcium resulted in the separation of EYFP and ECFP into monomeric fluorophores. In transiently transfected cell lines and dissociated hippocampal neurons, FRET was diminished by raising intracellular calcium levels with an ionophore or with glutamatergic agonists. Calpain inhibitors blocked these changes. Under control conditions, FRET levels in different dendritic spines of cultured neurons and in hippocampal slices were heterogeneous but showed robust decreases upon treatment with glutamatergic agonists. Immunostaining of cultured neurons with antibodies to a spectrin epitope produced by calpain-mediated digestion revealed an inverse correlation between the amount of FRET present at postsynaptic elements and the concentration of spectrin breakdown products. These results suggest that the FRET methodology identifies sites of synaptically induced calpain activity and that it may be useful in analyzing synapses undergoing changes in efficacy.
Figures
References
-
- Lynch G, Granger R. J Cognit Neurosci. 1992;4:189–199. - PubMed
-
- Roman F S, Chaillan F A, Soumireu-Mourat B. Brain Res. 1993;601:265–272. - PubMed
-
- Maren S. Trends Neurosci. 1999;22:561–567. - PubMed
-
- Malenka R C, Kauer J A, Zucker R S, Nicoll R A. Science. 1988;242:81–84. - PubMed
-
- Lynch G, Larson J, Kelso S, Barrionuevo G, Schottler F. Nature (London) 1983;305:719–721. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
