

Review
doi: 10.1128/JB.182.6.1459-1471.2000.
Signaling components in bacterial locomotion and sensory reception
Affiliations
- PMID: 10692349
- PMCID: PMC94441
- DOI: 10.1128/JB.182.6.1459-1471.2000
Item in Clipboard
Review
Signaling components in bacterial locomotion and sensory reception
J Bacteriol.
2000 Mar.
No abstract available
Figures
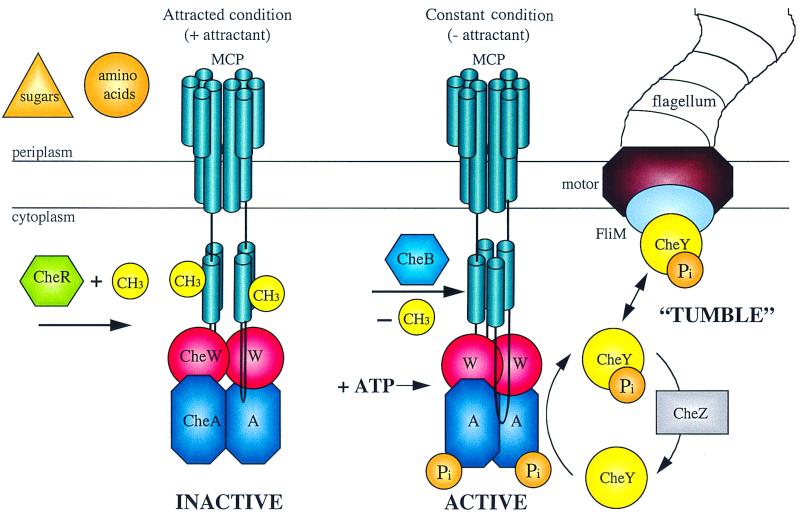
Schematic representation of the protein interactions and covalent modification during chemotaxis in enteric bacteria. Chemoeffector binding or release from the transmembrane MCP signal transducers results in a transmembrane conformational change that alters the activity of the CheA protein kinase and the susceptibility of the MCP protein to methylation or demethylation. Phosphorylated CheA can donate the phosphate to CheY, and P-CheY interacts with FliM in the flagellar basal body to influence the direction of flagellar rotation. (Figure provided by Jayna Ditta and Caroline Harwood.)
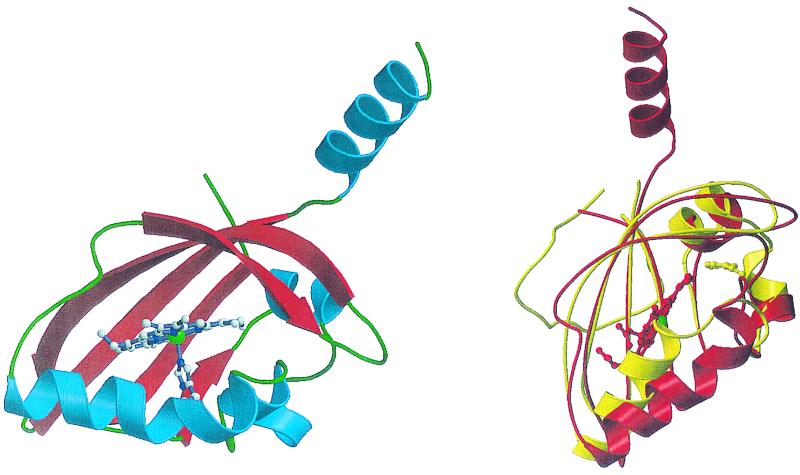
Structure of the heme-binding PAS domain from the FixL protein of Bradyrhizobium japonicum. (Left) A ribbons diagram with α-helices in blue and β-sheets in red. The helix pointing up leads to the HK domain. The heme moiety, with the heme iron in green, and its axial histidine ligand are shown. (Right) A comparison of the structures of the FixL heme-binding PAS domain in red and the PAS domain of the photoactive yellow protein PYP in yellow. The locations of the heme moiety of FixL, with the heme iron in green, and the p-hydroxycinnamate chromophore of PYP are indicated. Reprinted from reference with permission of the publisher. (Figure provided by M.-A. Gilles-Gonzalez.)
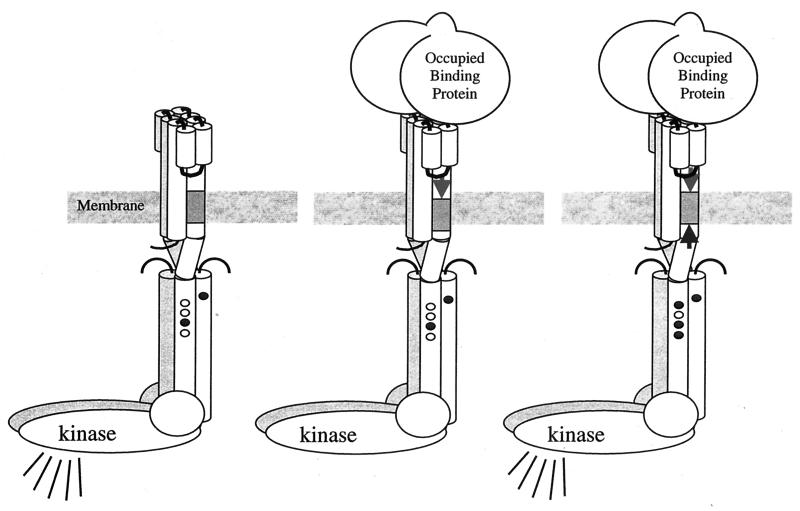
Helical sliding mediates transmembrane signaling and compensatory adaptation. Cartoon representation of the receptor-kinase complex shows sites of adaptational methylation (small ovals) in methylated (shaded) and unmethylated (white) states. (Left) In the absence of ligand, complexed kinase is active (symbolized by broadcast lines). (Middle) Binding of ligand (e.g., sugar-occupied, sugar-binding protein) induces sliding of TM2 toward cytoplasm (arrow pointing down), resulting in kinase inhibition (no broadcast lines). (Right) Increased adaptational methylation slides TM2 back toward the periplasm (arrow pointing up), and kinase activity is restored. (Figure provided by Bryan Beel and Gerald Hazelbauer.)
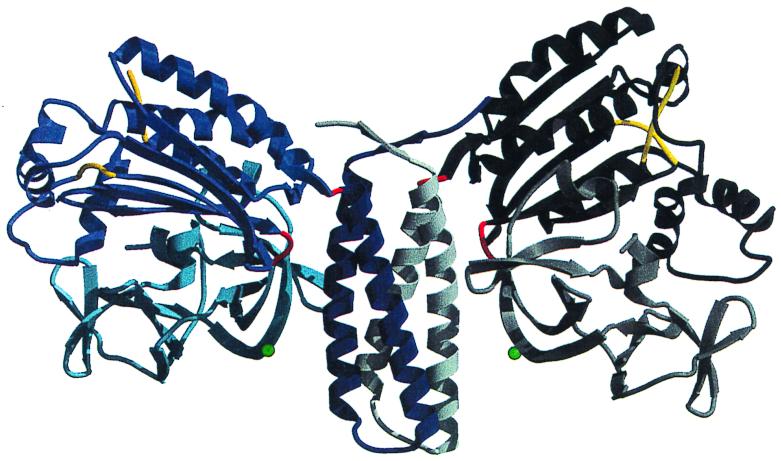
Crystal structure of the dimerization, ATP-binding, and regulatory domains of the histidine kinase CheA. CheA is the central component of the signaling pathway that controls bacterial chemotaxis. This is the first crystal structure of a histidine kinase. It reveals the modular composition of CheA, where the three functions of dimerization, ATP binding, and regulation are segregated to three separate domains. The domains are linked by hinges (in red) that allow movement. As a result, the interfaces between domains are different in the two subunits. Unexpectedly, the ATP-binding domain is different from other known kinases but resembles the ATP-binding domain of three ATPases, the topoisomerase gyrase B, the DNA repair enzyme MutL, and the chaperone Hsp90. Subunit A is shown in blue tones, whereas subunit B is shown in gray tones. The dimerization (middle of the figure) is a four-helix bundle where two helices are provided by each monomer (dark blue and white). The two ATP-binding domains (black and blue) are independent and are separated by 90 Å. Each regulatory domain (light gray and light blue) forms two β-barrels of topology reminiscent of SH3 domains. Reprinted from reference with permission of the publisher. (Figure provided by A. Bilwes.)
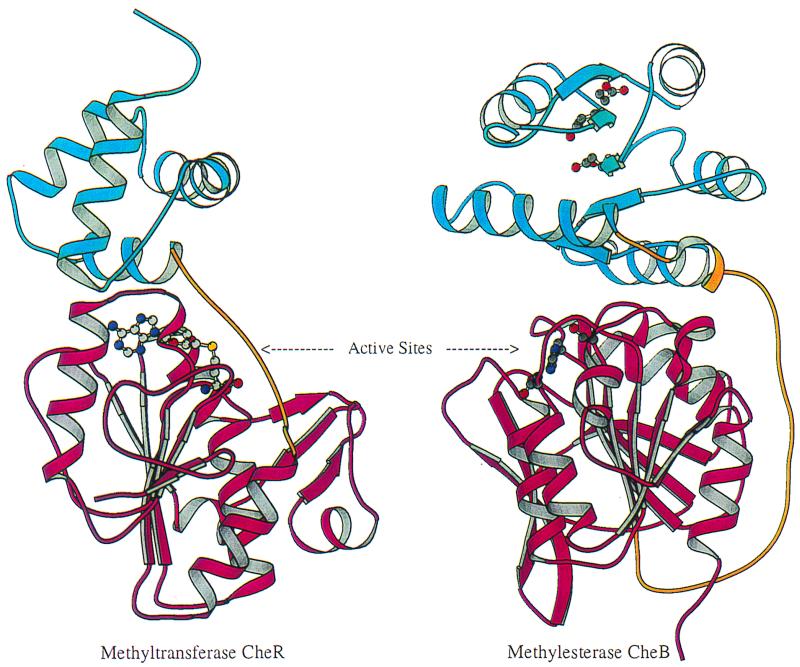
Structural comparison of receptor modification enzymes methyltransferase CheR and methylesterase CheB. Each protein contains two domains, an N-terminal domain (blue) and a C-terminal domain (magenta) connected by a linker (gold). The images are oriented to show the similar topologies of the C-terminal domains, the cofactor-binding domain of CheR, and the methylesterase catalytic domain of CheB. The S-adenosylmethionine cofactor in CheR and the active sites for phosphotransfer and methylesterase activities in CheB are depicted by ball-and-stick models (carbon, oxygen, nitrogen, and sulfur are shown in gray, red, blue, and yellow, respectively). Images were prepared using MOLSCRIPT (16). (Figure provided by Ann Stock.)
References
-
- Armitage J P, Schmitt R. Bacterial chemotaxis: Rhodobacter sphaeroides and Sinorhizobium meliloti—variations of a theme? Microbiology. 1997;143:3671–3682. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
