

Review
doi: 10.1093/emboj/19.5.800.
The importance of aquaporin water channel protein structures
Affiliations
- PMID: 10698922
- PMCID: PMC305620
- DOI: 10.1093/emboj/19.5.800
Item in Clipboard
Review
The importance of aquaporin water channel protein structures
EMBO J.
.
Abstract
The history of the water channel and recent structural and functional analyses of aquaporins are reviewed. These ubiquitous channels are important for bacteria, plants and animals, exhibit a pronounced sequence homology and share functional as well as structural similarities. Aquaporins allow water or small specific solutes to pass unhindered, but block the passage of ions to prevent dissipation of the transmembrane potential. Besides advances in structure determination, recent experiments suggest that many of these channels are regulated by pH variations, phosphorylation and binding of auxiliary proteins.
Figures
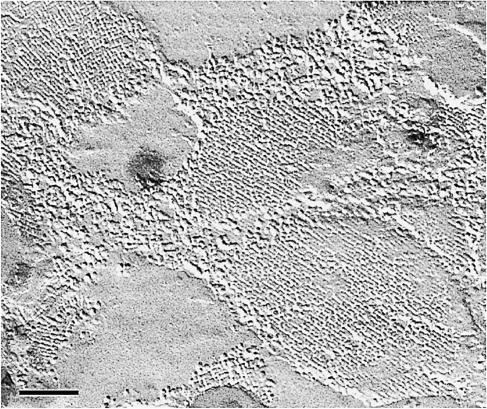
Fig. 1. Tetragonal arrays in lens fiber cell membranes are revealed by freeze–fracture techniques. The arrays are assembled from the major intrinsic protein, MIP (Kistler et al., 1980), the first member of the water channel protein family to be sequenced (Gorin et al., 1984) (by courtesy of J.Kistler, unpublished). Scale bar, 100 nm.
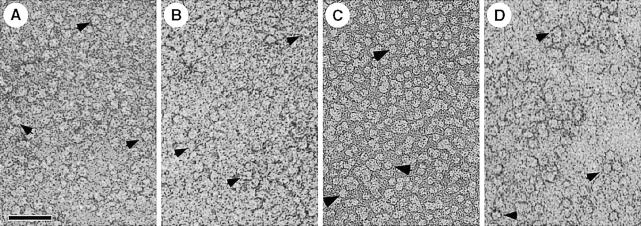
Fig. 2. All water channel proteins shown are isolated as tetramers (arrows) after solubilization of the respective membranes with octyl glucoside. (A) AQP1 from erythrocyte ghosts (Walz et al., 1994a). (B) AQP0 from ovine lens fiber cells (Hasler et al., 1998). (C) AqpZ from E.coli cells overexpressing this water channel (Ringler et al., 1999). (D) GlpF from E.coli cells overexpressing the glycerol facilitator (by courtesy of T.Braun, A.Philippsen and H.Stahlberg, unpublished). Scale bar, 50 nm.

Fig. 3. Projection maps of water channel proteins acquired by cryo-electron microscopy and calculated using the MRC software (Crowther et al., 1996). The maps reveal the structural similarity of the protein core that is embedded in the lipid bilayer and well conserved during sample preparation in spite of surface tension and interaction with the supporting carbon film. (A) The red cell water channel, AQP1, at 3.5 Å resolution. AQP1 packs into arrays with P4212 symmetry, housing two tetramers per unit cell of size 96 Å. (B) The lens fiber cell water channel, MIP (AQP0), at 5.7 Å resolution (D.Fotiadis, L.Hasler, D.J.Müller, H.Stahlberg, J.Kistler and A.Engel, manuscript submitted). AQP0 packs into P4 arrays with a single tetramer per unit cell of 64 Å side length. The area shown comprises two unit cells. (C) The bacterial water channel, AqpZ, at 8 Å resolution (Ringler et al., 1999). AqpZ is packed in an up-and-down orientation, as is AQP1, into unit cells of 94 Å width. All projections are viewed from the cytoplasmic side.
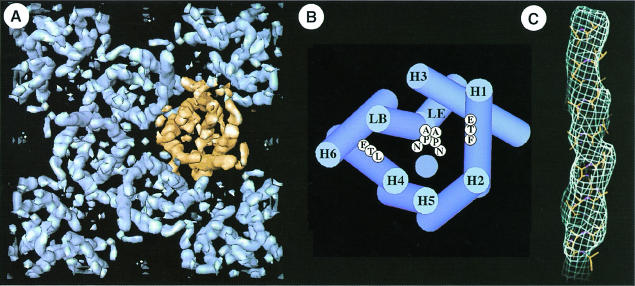
Fig. 4. The tetrameric arrangement of AQP1 and its molecular architecture is demonstrated three-dimensionally at 6 Å resolution (Walz et al., 1997). (A) Cytoplasmic view of one unit cell comprising the central tetramer and four monomers in the opposite orientation at the corners. Gaps within the tetramer indicate the monomer boundary. (B) The helix assignment derived from multiple sequence alignment (Heymann and Engel, 2000) is different from that proposed in Walz et al. (1997), but compatible with the 4.5 Å map established recently (Mitsuoka et al., 1999). The two NPA motifs are shown in the middle, and the hydrophobic residues on H1 (F) and H4 (L) are proposed to lie close to the channel (blue circle). (C) At 4.5 Å resolution, the helical nature of a membrane-spanning segment is revealed (Mitsuoka et al., 1999).
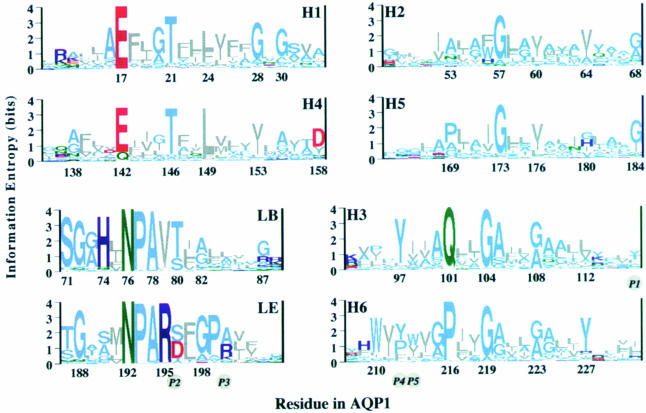
Fig. 5. Multiple alignment of 164 AQP/GLP sequences and subsequent phylogenetic analyses yielded 46 subtypes (Heymann and Engel, 1999). To identify critical residues, the 46 characteristic sequences were aligned and the conservation of each residue calculated (Heymann and Engel, 2000). Sequence logos, whose heights are a measure of conservation, are drawn with the residue numbers for AQP1. They reveal the conservation patterns of helices H1–H6 and the highly conserved loops, LB and LE. Helices are grouped in pairs according to the sequence similarity between the first and second half of the protein. The five positions (P1–P5) that were found by Froger et al. (1998) to be different between the AQP and GLP clusters are given in italics in circles. Colors: gray, hydrophobic; light blue, polar; green, amide; red, acidic; dark blue, basic.

Fig. 6. Surface topographies of water channel proteins are distinctly different, reflecting the differences in their sequences that are found mainly in the helix-connecting loops. Surface reliefs are acquired in buffer solution by atomic force microscopy. (A) Two-dimensional crystals assembled from AQP0 tetramers possess a P4 symmetry with one tetramer per unit cell of 64 Å width (Hasler et al., 1998). Four bilobed domains protrude by 14 Å from the extracellular surface of AQP0 (left). They are involved in a ‘tongue and groove’ interaction in junctions formed by two packed crystal layers (D.Fotiadis, L.Hasler, D.J.Müller, H.Stahlberg, J.Kistler and A.Engel, manuscript submitted). The cytoplasmic surface exhibits four domains of 8 Å in height surrounding a depression about the 4–fold axis (center). Carboxy-peptidase Y treatment on mica removes these domains and identifies them as carboxylic (D.Fotiadis, L.Hasler, D.J.Müller, H.Stahlberg, J.Kistler and A.Engel, manuscript submitted) (right). (B) AQP1 crystals have a P4212 symmetry and a corrugated extracellular surface with four protrusions of 12 Å in height (left). The cytosolic surface with four elongated peripheral and four small central domains produce a windmill-shaped structure with a height of 6 Å (center). Preliminary data from carboxypeptidase Y treatment on mica indicate that the C–terminus is located at a similar position as in AQP0 (D.J.Müller and A.Engel, unpublished data; right). (C) AqpZ has an extracellular surface with four elongated peripheral protrusions and four central protrusions of 7 Å in height (left). The poly-His tag used for purification produces a massive protrusion on the cytosolic surface (center). After trypsin cleavage, the cytosolic surface exhibits four small protrusions (right). The elongated extracellular protrusion in AqpZ has been identified as loop C based on its volume and flexibility (Scheuring et al., 1999). Correlation averages displayed comprise two tetramers and have a side length of ∼95 Å. Horizontal and vertical scaling are identical, but the gray level range is adapted to go from black to full white within each topograph.
References
-
- Borgnia M., Kozono, D., Calamita, G., Nielsen, S., Maloney, P.C. and Agre, P. (1999) Functional reconstitution and characterization of E.coli aquaporin-Z. J. Mol. Biol., 291, 1169–1179. - PubMed
-
- Cahalan K.L. and Hall, J.E. (1999) pH sensitivity of MIP-induced water permeability may play a role in regulating the intrinsic fluid circulation of the lens. Biophys. J., 76, A183.
-
- Cheng A., van Hoek, A.N., Yeager, M., Verkman, A.S. and Mitra, A.K. (1997) Three-dimensional organization of a human water channel. Nature, 387, 627–630. - PubMed
-
- Cooper G.J. and Boron, W.F. (1998) Effect of PCMBS on CO2 permeability of Xenopus oocytes expressing aquaporin 1 or its C189S mutant. Am. J. Physiol., 275, C1481–C1486. - PubMed
-
- Crowther R.A., Henderson, R. and Smith, J.M. (1996) MRC image processing programs. J. Struct. Biol., 116, 9–16. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
