

FADD/MORT1 regulates the pre-TCR checkpoint and can function as a tumour suppressor
- PMID: 10698935
- PMCID: PMC305633
- DOI: 10.1093/emboj/19.5.931
FADD/MORT1 regulates the pre-TCR checkpoint and can function as a tumour suppressor
Abstract
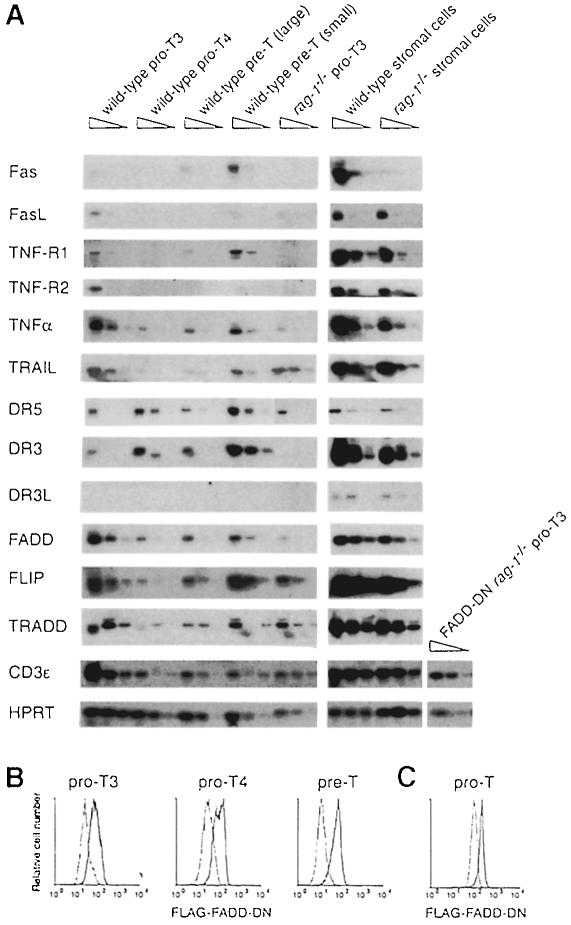
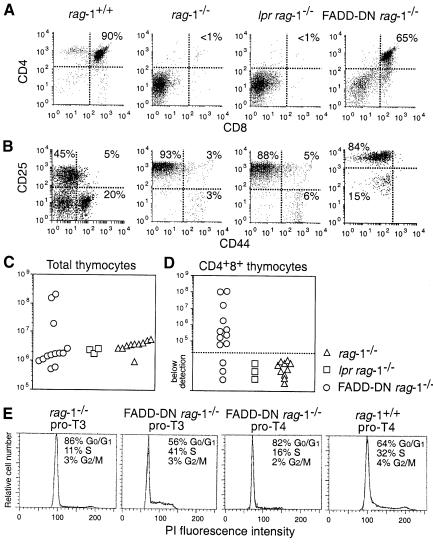
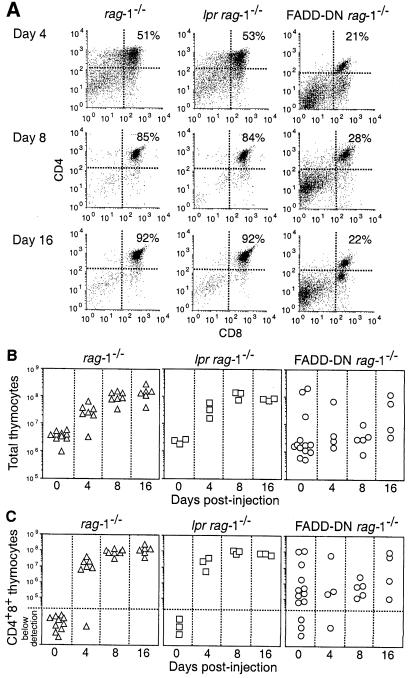
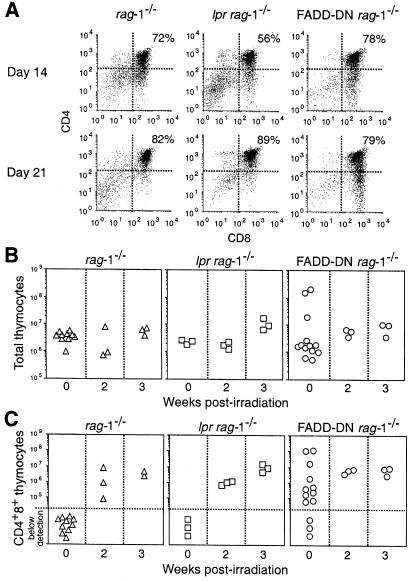
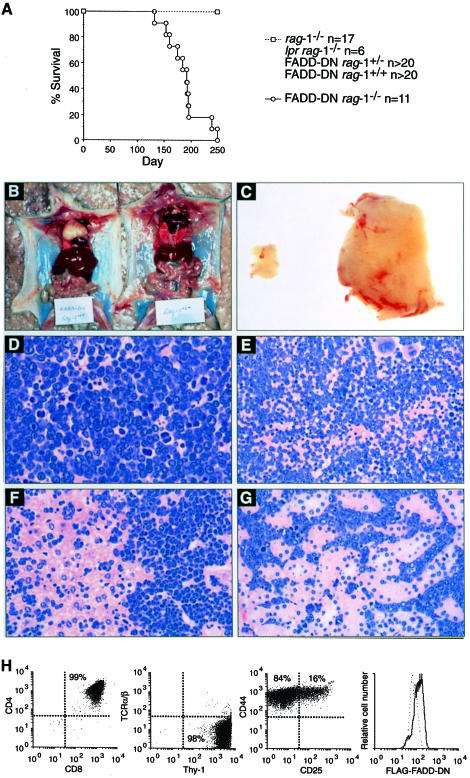
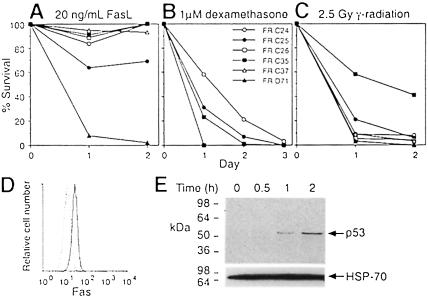
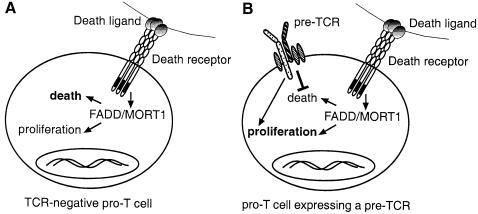
Productive rearrangement of the T-cell receptor (TCR) beta gene and signalling through the pre-TCR-CD3 complex are required for survival, proliferation and differentiation of T-cell progenitors (pro-T cells). Here we identify a role for death receptor signalling in early T-cell development using a dominant-negative mutant of the death receptor signal transducer FADD/MORT1 (FADD-DN). In rag-1(-/-) thymocytes, which are defective in antigen receptor gene rearrangement, FADD-DN bypassed the requirement for pre-TCR signalling, promoting pro-T-cell survival and differentiation to the more mature pre-T stage. Surprisingly, differentiation was not accompanied by the proliferation that occurs normally during transition to the pre-T stage. Consistent with a role for FADD/MORT1 in this cell division, FADD-DN rag-1(-/-) pro-T cells failed to proliferate in response to CD3epsilon ligation. Concomitant signalling through the pre-TCR and death receptors appears to trigger pro-T cell survival, proliferation and differentiation, whereas death receptor signalling in thymocytes that lack a pre-TCR induces apoptosis. Later in life all FADD-DN rag-1(-/-) mice developed thymic lymphoma, indicating that FADD/MORT1 can act as a tumour suppressor.
Figures
Similar articles
-
Fas-mediated apoptosis and activation-induced T-cell proliferation are defective in mice lacking FADD/Mort1.Nature. 1998 Mar 19;392(6673):296-300. doi: 10.1038/32681. Nature. 1998. PMID: 9521326
-
A dominant interfering mutant of FADD/MORT1 enhances deletion of autoreactive thymocytes and inhibits proliferation of mature T lymphocytes.EMBO J. 1998 Feb 2;17(3):706-18. doi: 10.1093/emboj/17.3.706. EMBO J. 1998. PMID: 9450996 Free PMC article.
-
p53-dependent impairment of T-cell proliferation in FADD dominant-negative transgenic mice.Curr Biol. 1998 Apr 9;8(8):467-70. doi: 10.1016/s0960-9822(98)70182-4. Curr Biol. 1998. PMID: 9550704
-
FADD/MORT1, a signal transducer that can promote cell death or cell growth.Int J Biochem Cell Biol. 1999 May;31(5):533-7. doi: 10.1016/s1357-2725(99)00003-5. Int J Biochem Cell Biol. 1999. PMID: 10399313 Review.
-
Regulation of thymocyte differentiation: pre-TCR signals and beta-selection.Semin Immunol. 2002 Oct;14(5):311-23. doi: 10.1016/s1044-5323(02)00064-7. Semin Immunol. 2002. PMID: 12220932 Review.
Cited by
-
Cellular FLICE-inhibitory protein is required for T cell survival and cycling.J Exp Med. 2005 Aug 1;202(3):405-13. doi: 10.1084/jem.20050118. Epub 2005 Jul 25. J Exp Med. 2005. PMID: 16043518 Free PMC article.
-
Dominant Negative FADD/MORT1 Inhibits the Development of Intestinal Intraepithelial Lymphocytes With a Marked Defect on CD8αα+TCRγδ+ T Cells.Front Immunol. 2018 Sep 10;9:2038. doi: 10.3389/fimmu.2018.02038. eCollection 2018. Front Immunol. 2018. PMID: 30250469 Free PMC article.
-
Disruption of pre-TCR expression accelerates lymphomagenesis in E2A-deficient mice.Proc Natl Acad Sci U S A. 2002 Aug 20;99(17):11322-7. doi: 10.1073/pnas.162373999. Epub 2002 Aug 9. Proc Natl Acad Sci U S A. 2002. PMID: 12172006 Free PMC article.
-
DR3 regulates negative selection during thymocyte development.Mol Cell Biol. 2001 May;21(10):3451-61. doi: 10.1128/MCB.21.10.3451-3461.2001. Mol Cell Biol. 2001. PMID: 11313471 Free PMC article.
-
The GTPase rho controls a p53-dependent survival checkpoint during thymopoiesis.J Exp Med. 2000 Jul 3;192(1):77-85. doi: 10.1084/jem.192.1.77. J Exp Med. 2000. PMID: 10880528 Free PMC article.
References
-
- Akashi K., Kondo, M., von Freeden-Jeffry, U., Murray, R. and Weissman, I.L. (1997) Bcl-2 rescues T lymphopoiesis in interleukin-7 receptor-deficient mice. Cell, 89, 1033–1041. - PubMed
-
- Ashkenazi A. and Dixit, V.M. (1998) Death receptors: signaling and modulation. Science, 281, 1305–1308. - PubMed
-
- Boehme S.A. and Lenardo, M.J. (1996) TCR-mediated death of mature T lymphocytes occurs in the absence of p53. J. Immunol., 156, 4075–4078. - PubMed
-
- Bogue M.A., Zhu, C., Aguilar-Cordova, E., Donehower, L.A. and Roth, D.B. (1996) p53 is required for both radiation-induced differentiation and rescue of V(D)J rearrangement in scid mouse thymocytes. Genes Dev., 10, 553–565. - PubMed
-
- Boldin M.P., Varfolomeev, E.E., Pancer, Z., Mett, I.L., Camonis, J.H. and Wallach, D. (1995) A novel protein that interacts with the death domain of Fas/APO1 contains a sequence motif related to the death domain. J. Biol. Chem., 270, 7795–7798. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
