

UV damage causes uncontrolled DNA breakage in cells from patients with combined features of XP-D and Cockayne syndrome
- PMID: 10698956
- PMCID: PMC305654
- DOI: 10.1093/emboj/19.5.1157
UV damage causes uncontrolled DNA breakage in cells from patients with combined features of XP-D and Cockayne syndrome
Abstract
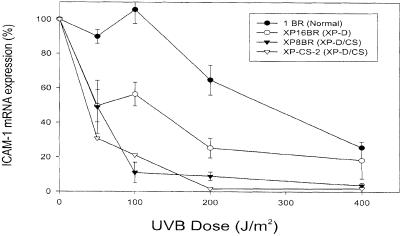
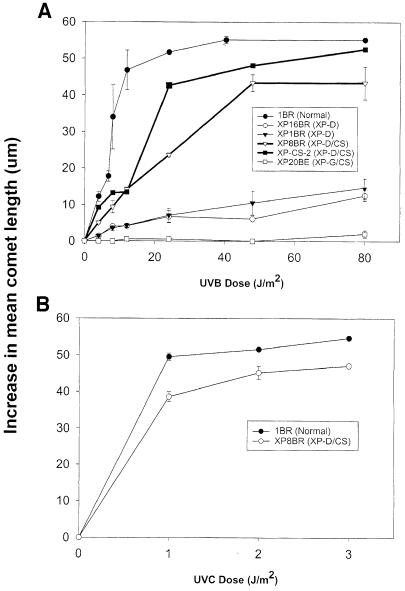
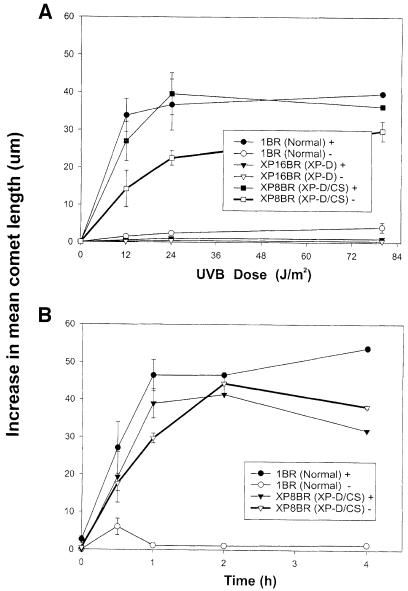
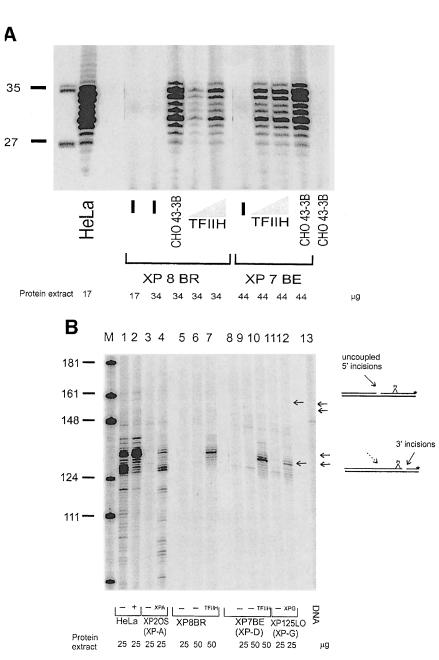
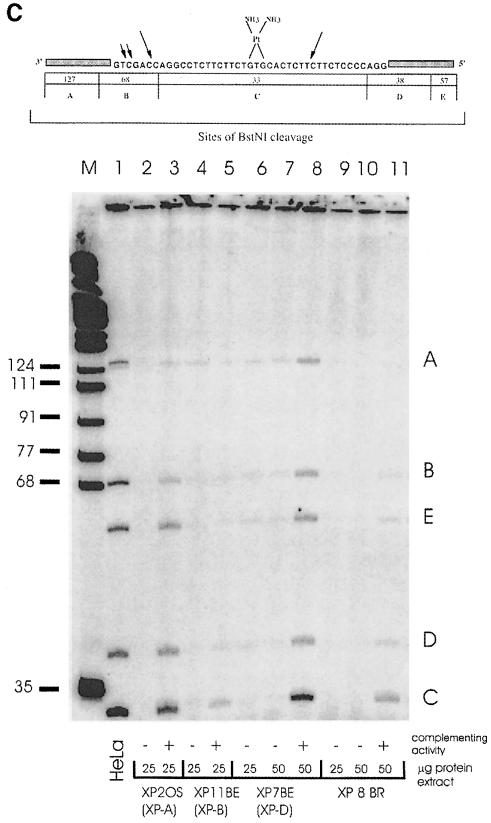
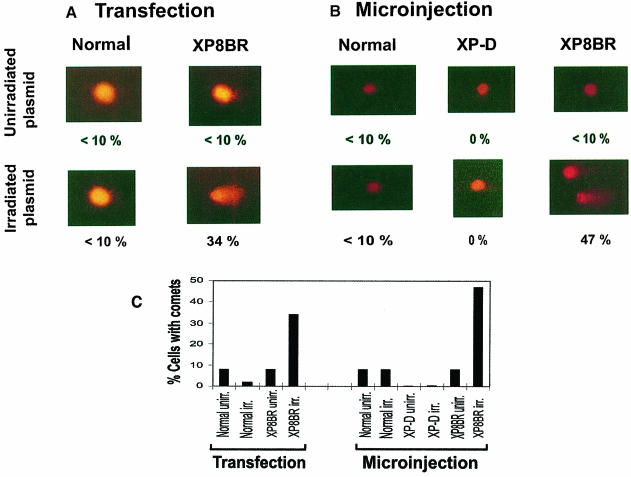
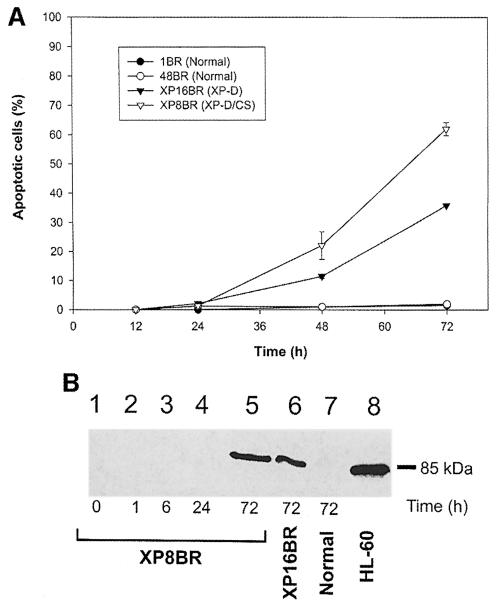
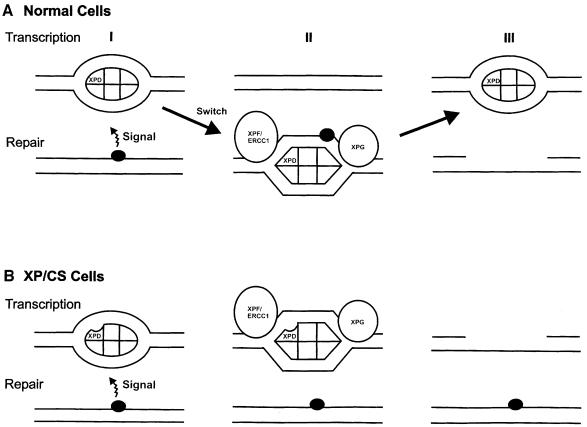
Nucleotide excision repair (NER) removes damage from DNA in a tightly regulated multiprotein process. Defects in NER result in three different human disorders, xeroderma pigmentosum (XP), trichothiodystrophy (TTD) and Cockayne syndrome (CS). Two cases with the combined features of XP and CS have been assigned to the XP-D complementation group. Despite their extreme UV sensitivity, these cells appeared to incise their DNA as efficiently as normal cells in response to UV damage. These incisions were, however, uncoupled from the rest of the repair process. Using cell-free extracts, we were unable to detect any incision activity in the neighbourhood of the damage. When irradiated plasmids were introduced into unirradiated XP-D/CS cells, the ectopically introduced damage triggered the induction of breaks in the undamaged genomic DNA. XP-D/CS cells thus have a unique response to sensing UV damage, which results in the introduction of breaks into the DNA at sites distant from the damage. We propose that it is these spurious breaks that are responsible for the extreme UV sensitivity of these cells.
Figures
References
-
- Aboussekhra A., et al. (1995) Mammalian DNA nucleotide excision repair reconstituted with purified components. Cell, 80, 859–868. - PubMed
-
- Berneburg M., Clingen, P.H., Harcourt, S.A., Lowe, J.E., Taylor, E.M., Green, M.H.L., Krutmann, J., Arlett, C.F. and Lehmann, A.R. (2000) The cancer-free phenotype in trichothiodystrophy is unrelated to its repair defect. Cancer Res., 60, 431–438. - PubMed
-
- Botta E., Nardo, T., Broughton, B.C., Marinoni, S., Lehmann, A.R. and Stefanini, M. (1998) Analysis of mutations in the XPD gene in Italian patients with trichothiodystrophy: site of mutation correlates with repair deficiency but gene dosage appears to determine clinical severity. Am. J. Hum. Genet., 63, 190–196. - PMC - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Research Materials
