

Nuclear export of MAP kinase (ERK) involves a MAP kinase kinase (MEK)-dependent active transport mechanism
- PMID: 10704436
- PMCID: PMC2174544
- DOI: 10.1083/jcb.148.5.849
Nuclear export of MAP kinase (ERK) involves a MAP kinase kinase (MEK)-dependent active transport mechanism
Erratum in
- J Cell Biol 2000 May 1;149(3):754
Abstract
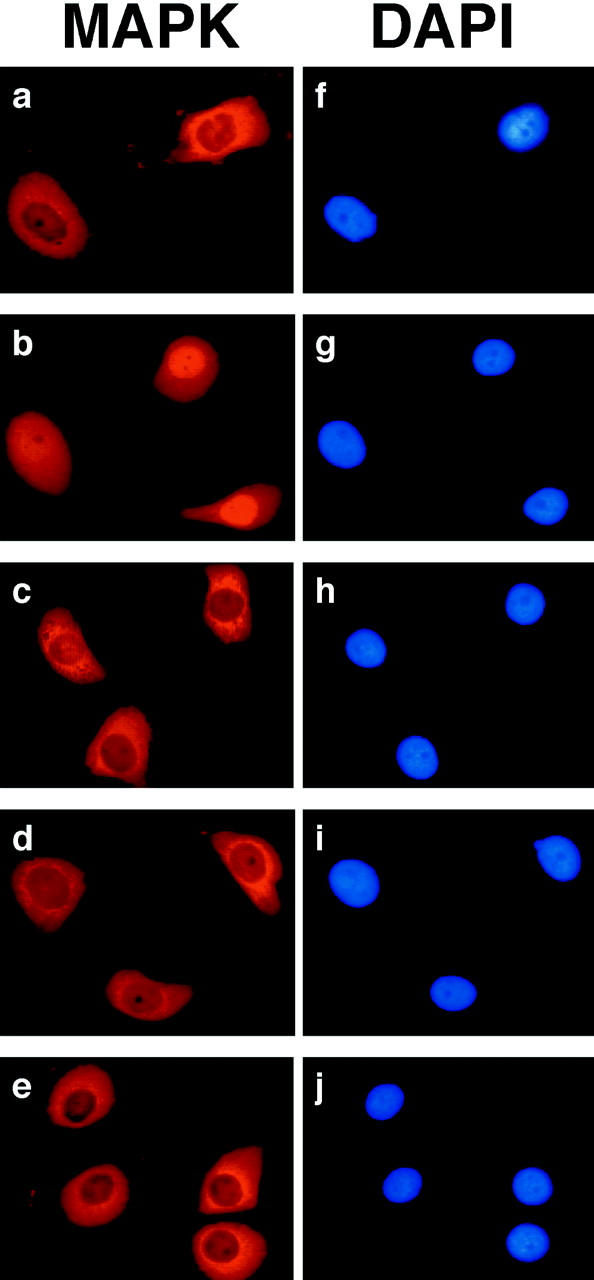
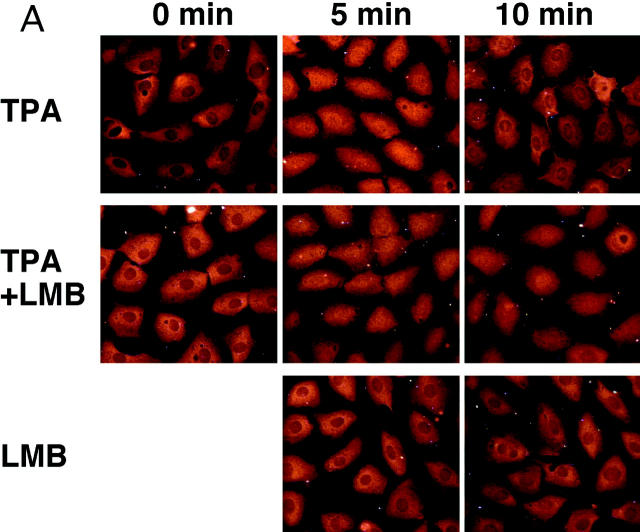
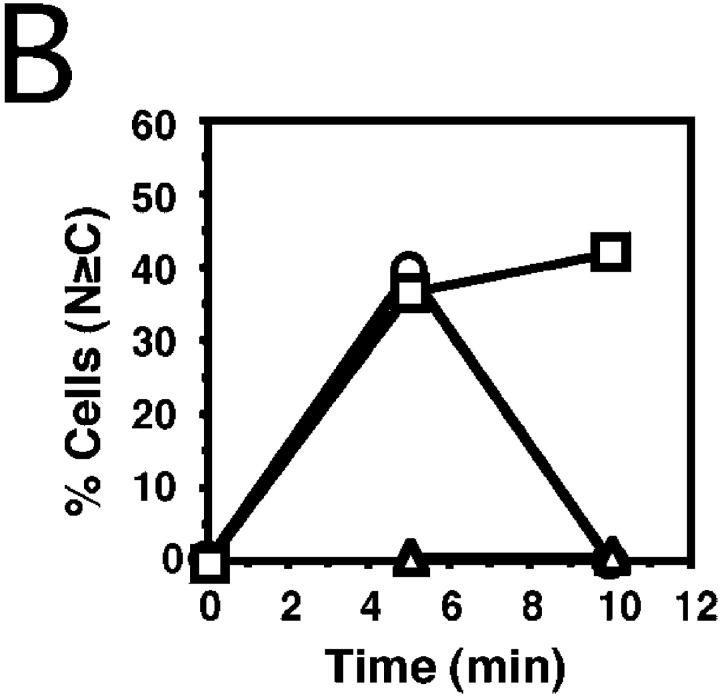
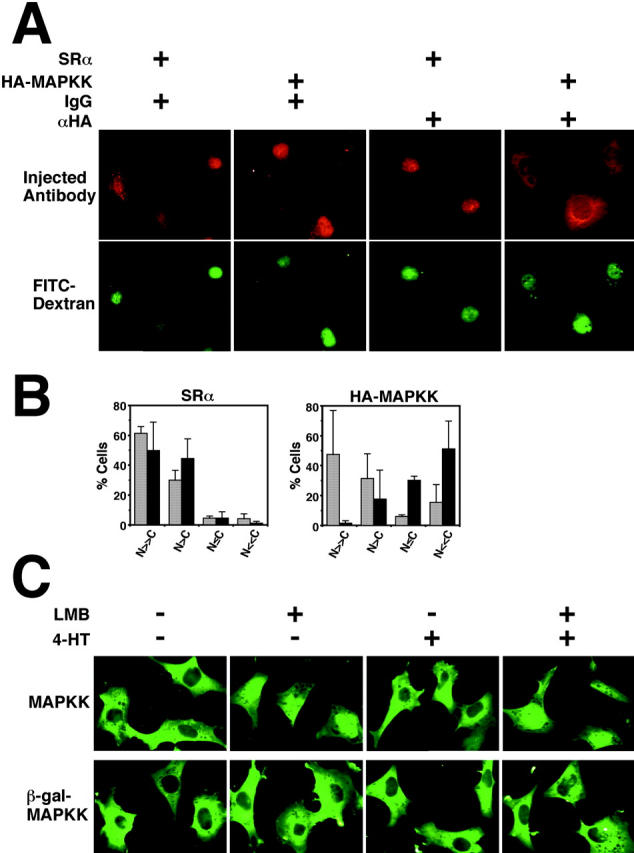
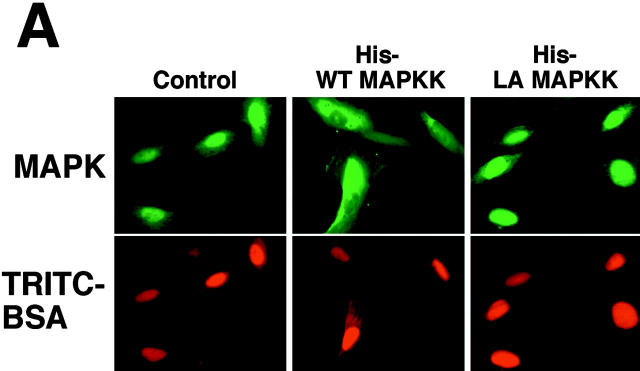
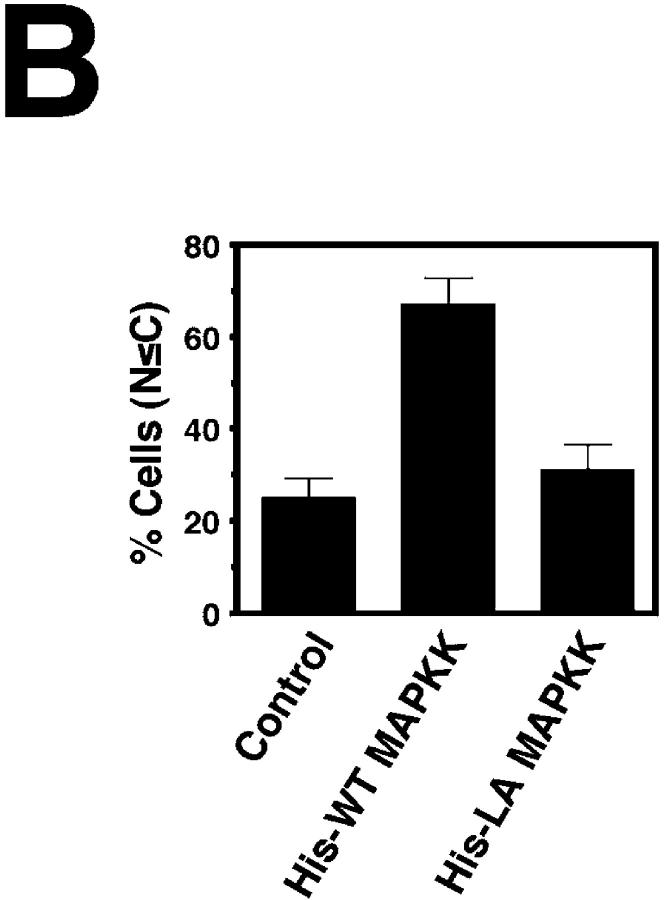
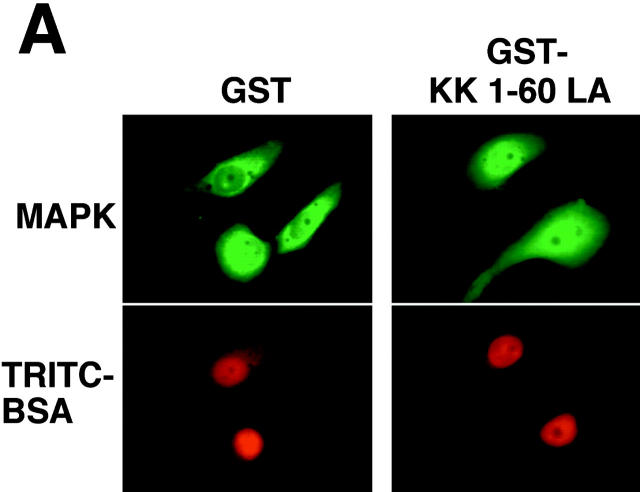
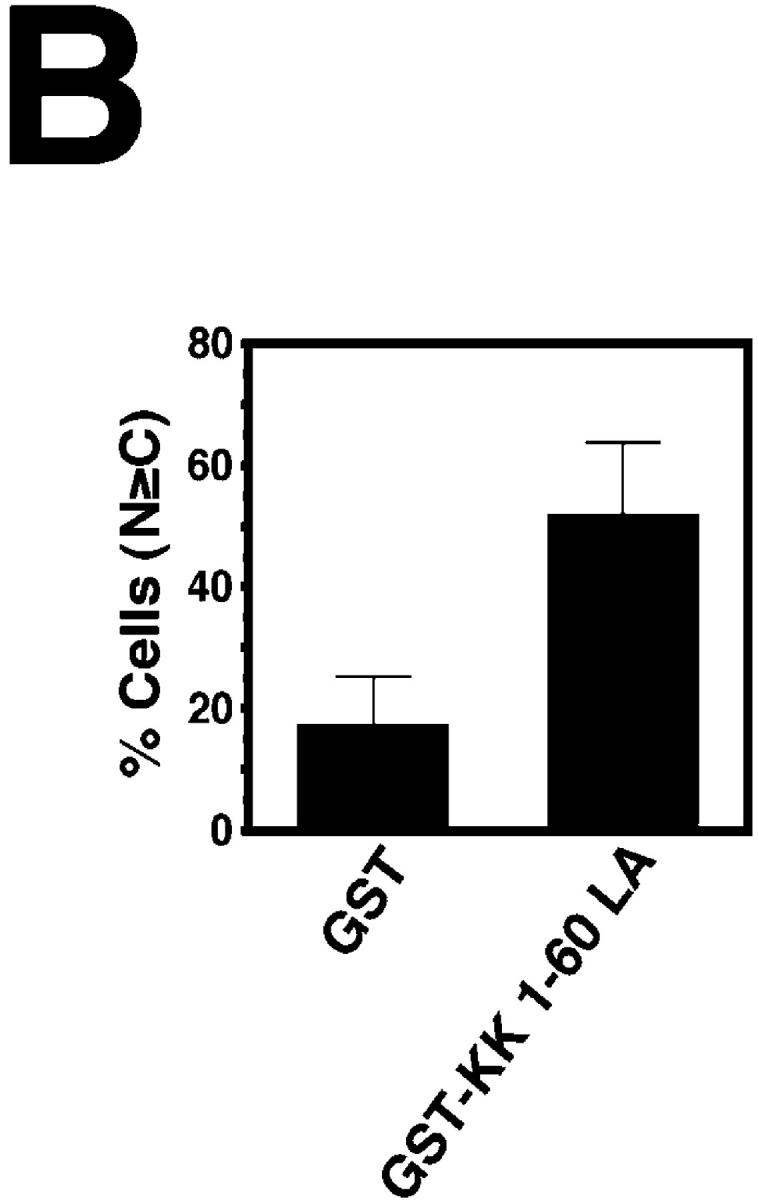
In response to extracellular stimuli, mitogen-activated protein kinase (MAPK, also known as ERK), which localizes to the cytoplasm in quiescent cells, translocates to the nucleus and then relocalizes to the cytoplasm again. The relocalization of nuclear MAPK to the cytoplasm was not inhibited by cycloheximide, confirming that the relocalization is achieved by nuclear export, but not synthesis, of MAPK. The nuclear export of MAPK was inhibited by leptomycin B (LMB), a specific inhibitor of the nuclear export signal (NES)-dependent transport. We have then shown that MAP kinase kinase (MAPKK, also known as MEK), which mostly localizes to the cytoplasm because of its having NES, is able to shuttle between the cytoplasm and the nucleus constantly. MAPK, when injected into the nucleus, was rapidly exported from the nucleus by coinjected wild-type MAPKK, but not by the NES-disrupted MAPKK. In addition, injection of the fragment corresponding to the MAPK-binding site of MAPKK into the nucleus, which would disrupt the binding of MAPK to MAPKK in the nucleus, significantly inhibited the nuclear export of endogenous MAPK. Taken together, these results suggest that the relocalization of nuclear MAPK to the cytoplasm involves a MAPKK-dependent, active transport mechanism.
Figures
References
-
- Bardwell L., Cook J.G., Chang E.C., Cairns B.R., Thorner J. Signaling in the yeast pheromone response pathwayspecific and high-affinity interaction of the mitogen-activated protein (MAP) kinases Kss1 and Fus3 with the upstream MAP kinase kinase Ste7. Mol. Cell. Biol. 1996;16:3637–3650. - PMC - PubMed
-
- Davis R.J. The mitogen-activated protein kinase signal transduction pathway. J. Biol. Chem. 1993;268:14553–14556. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
