

MAD3 encodes a novel component of the spindle checkpoint which interacts with Bub3p, Cdc20p, and Mad2p
- PMID: 10704439
- PMCID: PMC2174553
- DOI: 10.1083/jcb.148.5.871
MAD3 encodes a novel component of the spindle checkpoint which interacts with Bub3p, Cdc20p, and Mad2p
Abstract
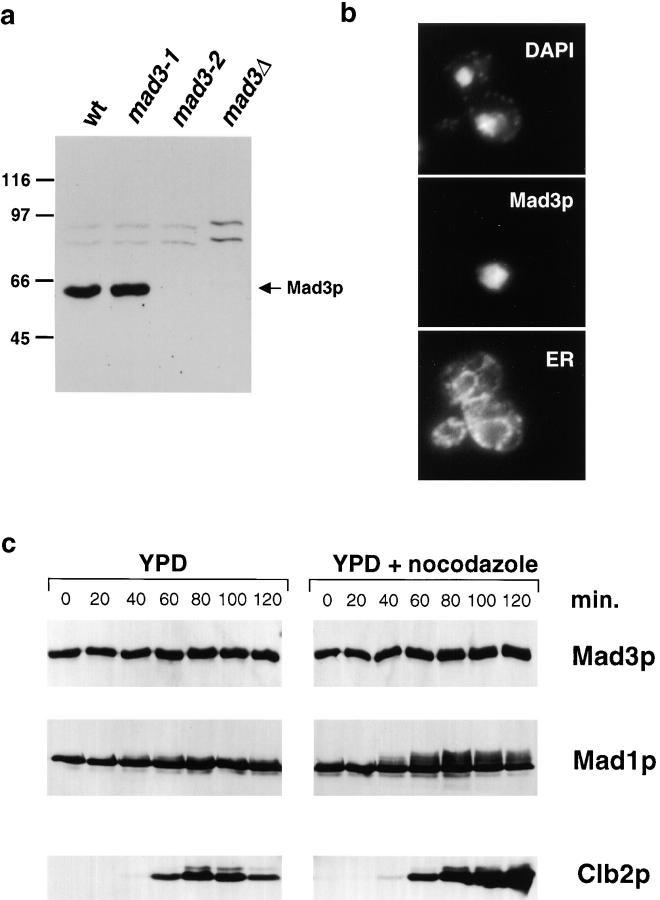
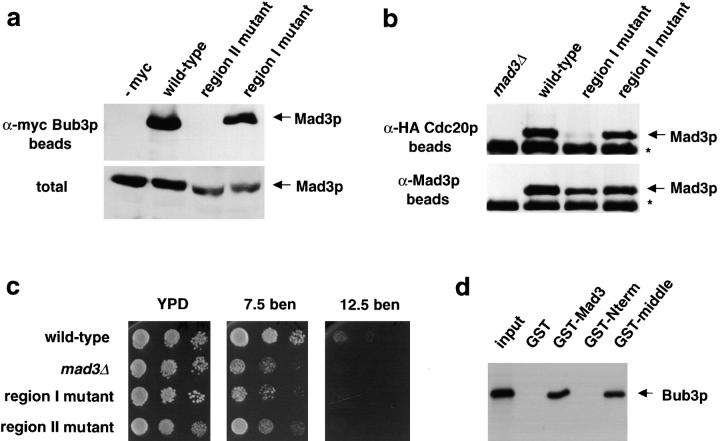
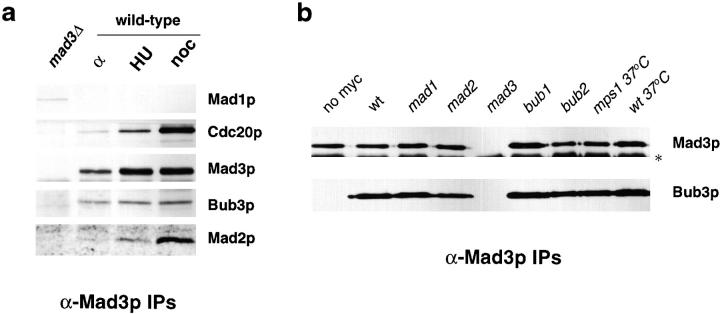
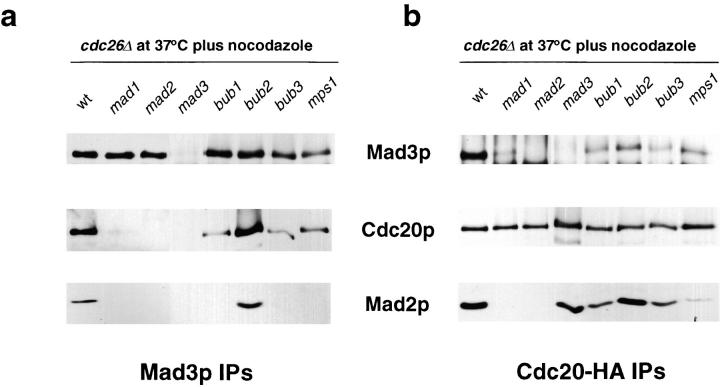
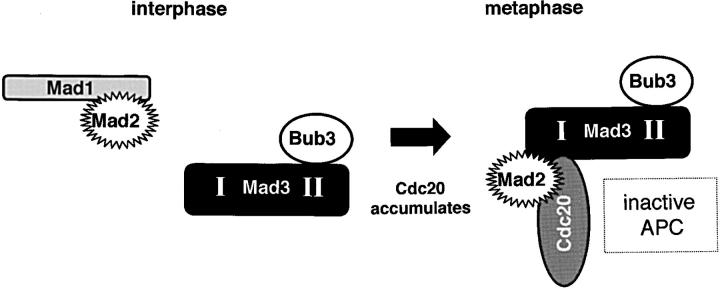
We show that MAD3 encodes a novel 58-kD nuclear protein which is not essential for viability, but is an integral component of the spindle checkpoint in budding yeast. Sequence analysis reveals two regions of Mad3p that are 46 and 47% identical to sequences in the NH(2)-terminal region of the budding yeast Bub1 protein kinase. Bub1p is known to bind Bub3p (Roberts et al. 1994) and we use two-hybrid assays and coimmunoprecipitation experiments to show that Mad3p can also bind to Bub3p. In addition, we find that Mad3p interacts with Mad2p and the cell cycle regulator Cdc20p. We show that the two regions of homology between Mad3p and Bub1p are crucial for these interactions and identify loss of function mutations within each domain of Mad3p. We discuss roles for Mad3p and its interactions with other spindle checkpoint proteins and with Cdc20p, the target of the checkpoint.
Figures

References
-
- Cahill D.P., Lengauer C., Yu J., Riggins G.J., Willson J.K.V., Markowitz S.D., Kinzler K.W., Vogelstein B. Mutations of mitotic checkpoint genes in human cancers. Nature. 1998;392:300–303. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
