

Deficiency of the stress kinase p38alpha results in embryonic lethality: characterization of the kinase dependence of stress responses of enzyme-deficient embryonic stem cells
- PMID: 10704466
- PMCID: PMC2195860
- DOI: 10.1084/jem.191.5.859
Deficiency of the stress kinase p38alpha results in embryonic lethality: characterization of the kinase dependence of stress responses of enzyme-deficient embryonic stem cells
Abstract
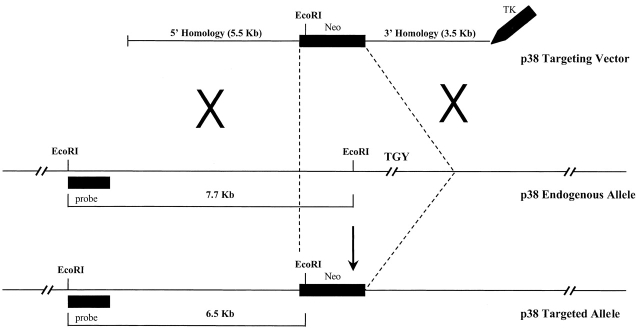
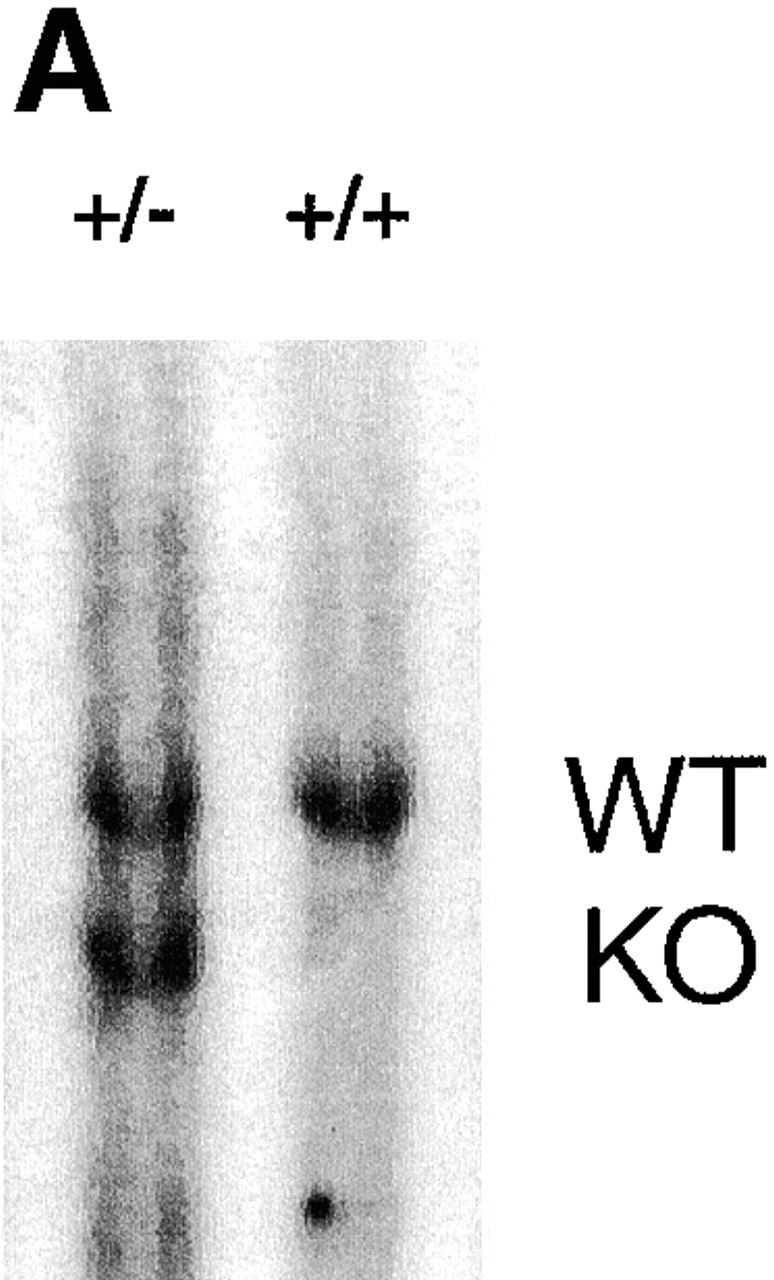
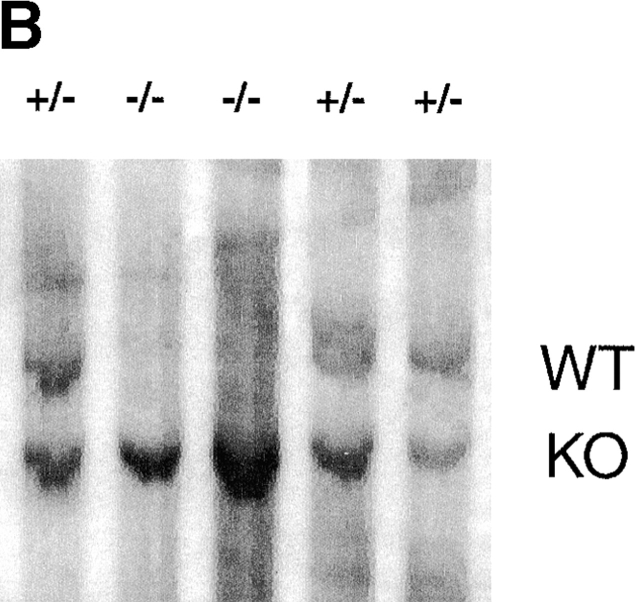
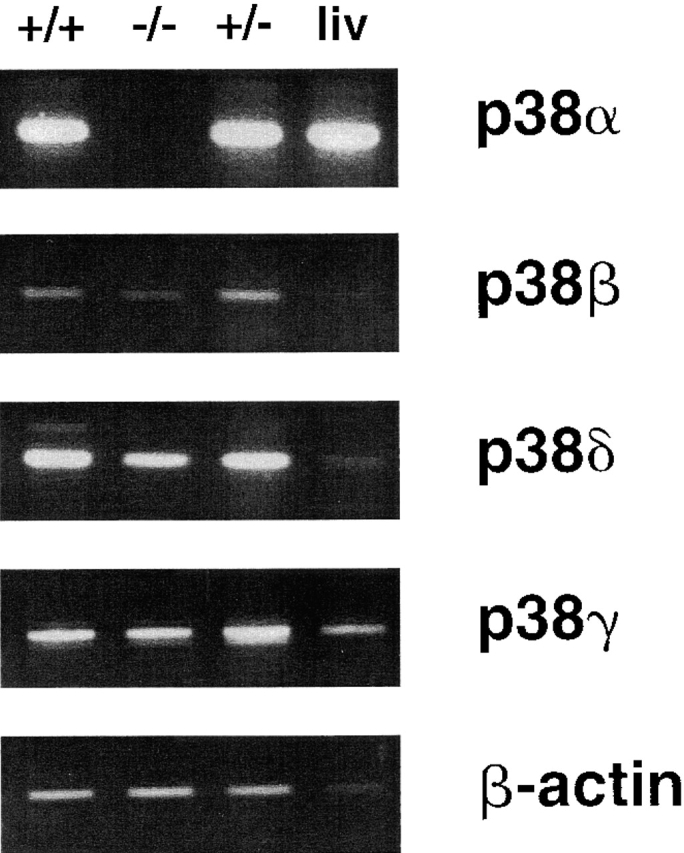
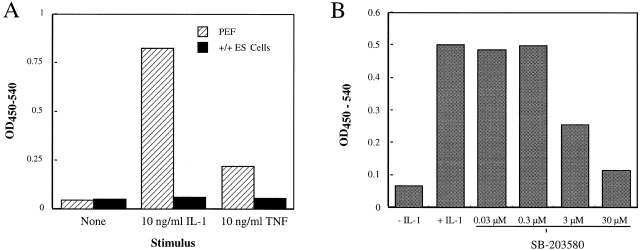
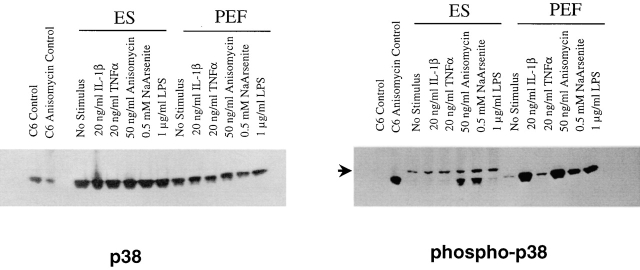
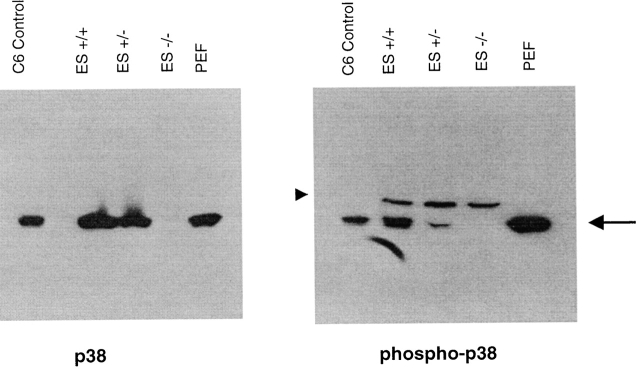
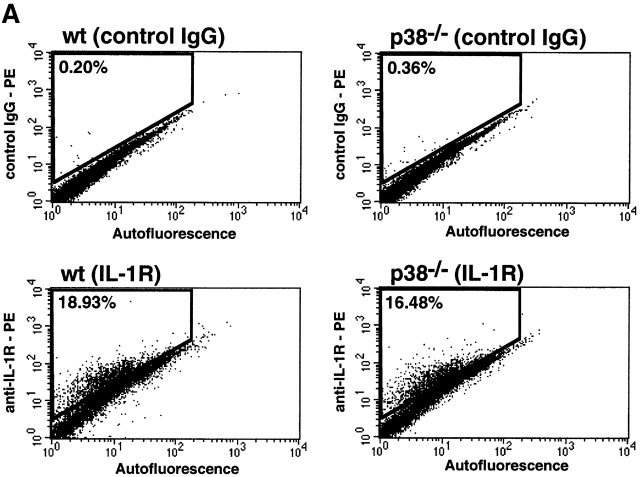
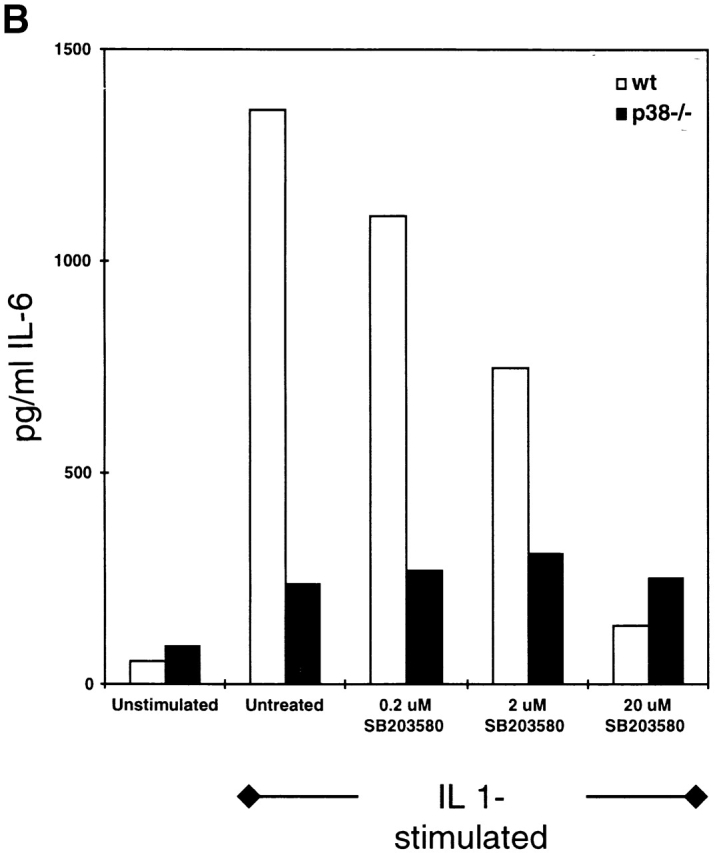
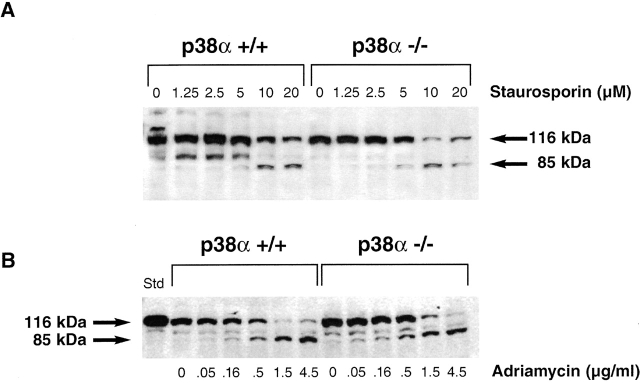
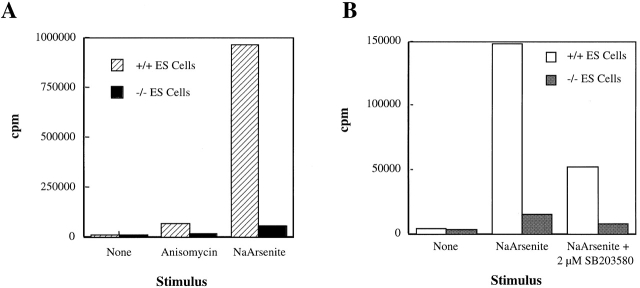
The mitogen-activated protein (MAP) kinase p38 is a key component of stress response pathways and the target of cytokine-suppressing antiinflammatory drugs (CSAIDs). A genetic approach was employed to inactivate the gene encoding one p38 isoform, p38alpha. Mice null for the p38alpha allele die during embryonic development. p38alpha(1/)- embryonic stem (ES) cells grown in the presence of high neomycin concentrations demonstrated conversion of the wild-type allele to a targeted allele. p38alpha(-/)- ES cells lacked p38alpha protein and failed to activate MAP kinase-activated protein (MAPKAP) kinase 2 in response to chemical stress inducers. In contrast, p38alpha(1/+) ES cells and primary embryonic fibroblasts responded to stress stimuli and phosphorylated p38alpha, and activated MAPKAP kinase 2. After in vitro differentiation, both wild-type and p38alpha(-/)- ES cells yielded cells that expressed the interleukin 1 receptor (IL-1R). p38alpha(1/+) but not p38alpha(-/)- IL-1R-positive cells responded to IL-1 activation to produce IL-6. Comparison of chemical-induced apoptosis processes revealed no significant difference between the p38alpha(1/+) and p38alpha(-/)- ES cells. Therefore, these studies demonstrate that p38alpha is a major upstream activator of MAPKAP kinase 2 and a key component of the IL-1 signaling pathway. However, p38alpha does not serve an indispensable role in apoptosis.
Figures
References
-
- Karin M. Mitogen-activated protein kinase cascades as regulators of stress responses. Ann. NY Acad. Sci. 1998;851:139–146. - PubMed
-
- Kyriakis J.M., Auruch J. Sounding the alarmprotein kinase cascades activated by stress and inflammation. J. Biol. Chem. 1996;271:24313–24316. - PubMed
-
- Ludwig S., Hoffmeyer A., Goebeler M., Kilian K., Hafner H., Neufeld B., Han J., Rap U.R. The stress inducer arsenite activates mitogen-activated protein kinases extracellular signal-regulated kinases 1 and 2 via a MAPK kinase 6/p38-dependent pathway. J. Biol. Chem. 1998;273:1917–1922. - PubMed
-
- Rouse J., Cohen P., Trigon S., Morange M., Alonso-Liamazares A., Zamanillo D., Hunt T., Nebreda A.R. A novel kinase cascade triggered by stress and heat shock that stimulates MAPKAP kinase-2 and phosphorylation of the small heat shock proteins. Cell. 1994;78:1027–1037. - PubMed
-
- Raingeaud J., Gupta S., Rogers J. S., Dickens M., Han J., Ulevitch R.J., Davis R.J. Pro-inflammatory cytokines and environmental stress cause p38 mitogen-activated protein kinase activation by dual phosphorylation on tyrosine and threonine. J. Biol. Chem. 1995;270:7420–7426. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
