

Genetics of mosquito vector competence
- PMID: 10704476
- PMCID: PMC98988
- DOI: 10.1128/MMBR.64.1.115-137.2000
Genetics of mosquito vector competence
Abstract
Mosquito-borne diseases are responsible for significant human morbidity and mortality throughout the world. Efforts to control mosquito-borne diseases have been impeded, in part, by the development of drug-resistant parasites, insecticide-resistant mosquitoes, and environmental concerns over the application of insecticides. Therefore, there is a need to develop novel disease control strategies that can complement or replace existing control methods. One such strategy is to generate pathogen-resistant mosquitoes from those that are susceptible. To this end, efforts have focused on isolating and characterizing genes that influence mosquito vector competence. It has been known for over 70 years that there is a genetic basis for the susceptibility of mosquitoes to parasites, but until the advent of powerful molecular biological tools and protocols, it was difficult to assess the interactions of pathogens with their host tissues within the mosquito at a molecular level. Moreover, it has been only recently that the molecular mechanisms responsible for pathogen destruction, such as melanotic encapsulation and immune peptide production, have been investigated. The molecular characterization of genes that influence vector competence is becoming routine, and with the development of the Sindbis virus transducing system, potential antipathogen genes now can be introduced into the mosquito and their effect on parasite development can be assessed in vivo. With the recent successes in the field of mosquito germ line transformation, it seems likely that the generation of a pathogen-resistant mosquito population from a susceptible population soon will become a reality.
Figures
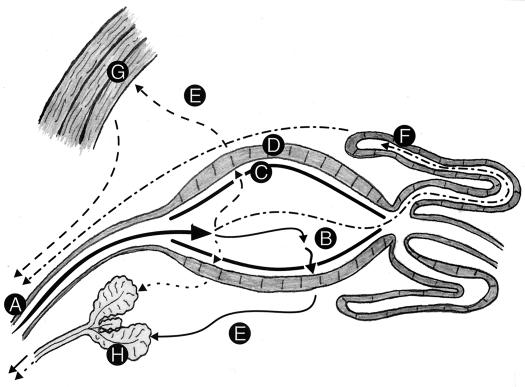
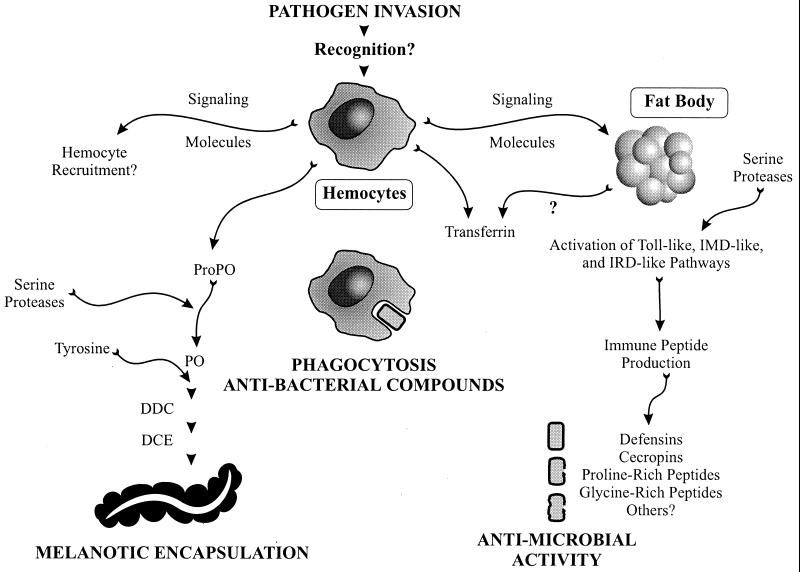
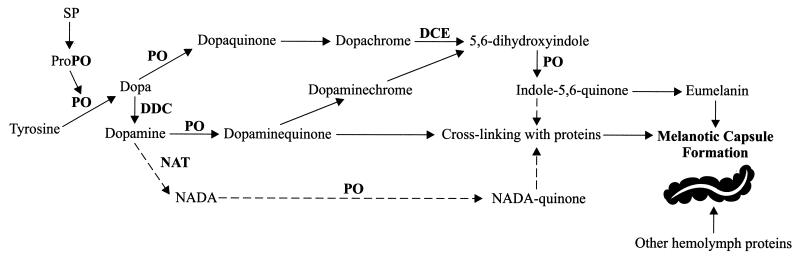

References
-
- Barillas-Mury C, Graf R, Hagedorn H H, Wells M A. cDNA and deduced amino acid sequence of a blood meal-induced trypsin from the mosquito, Aedes aegypti. Insect Biochem. 1991;21:825–831.
-
- Barreau C, Conrad J, Fischer E, Lujan H D, Vernick K D. Identification of surface molecules on salivary glands of the mosquito, Aedes aegypti, by a panel of monoclonal antibodies. Insect Biochem Mol Biol. 1999;29:515–526. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
