

Spectral-temporal receptive fields of nonlinear auditory neurons obtained using natural sounds
- PMID: 10704507
- PMCID: PMC6772498
- DOI: 10.1523/JNEUROSCI.20-06-02315.2000
Spectral-temporal receptive fields of nonlinear auditory neurons obtained using natural sounds
Abstract
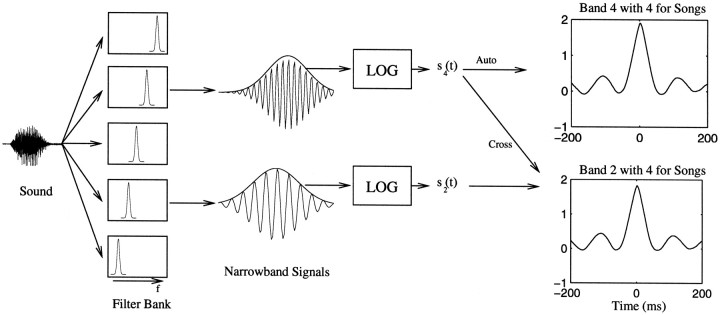
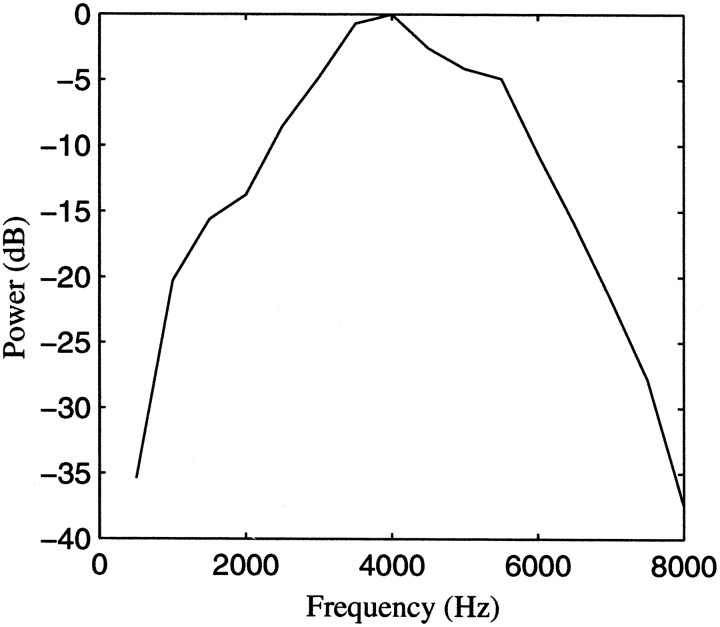
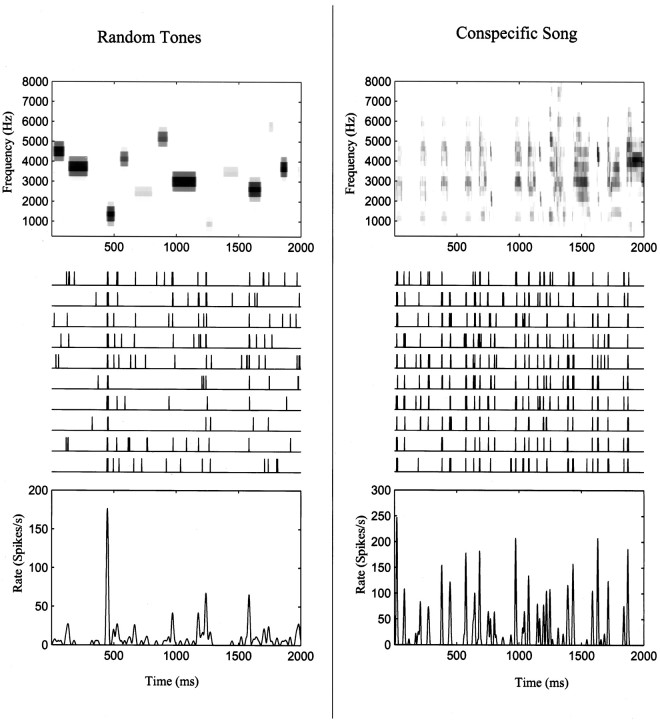
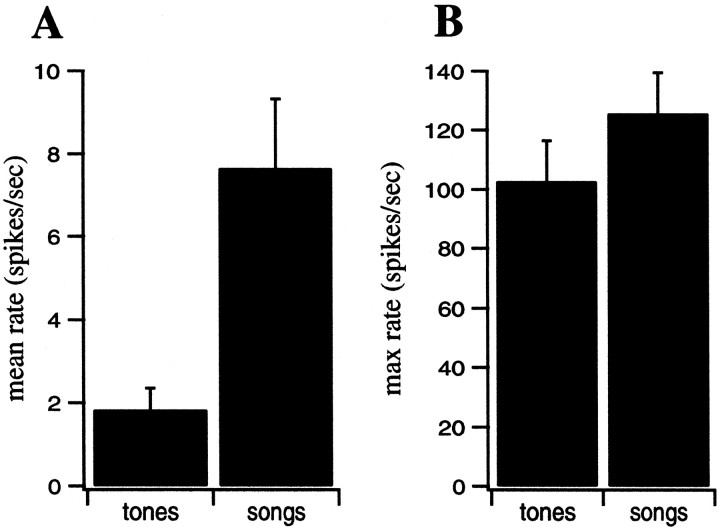
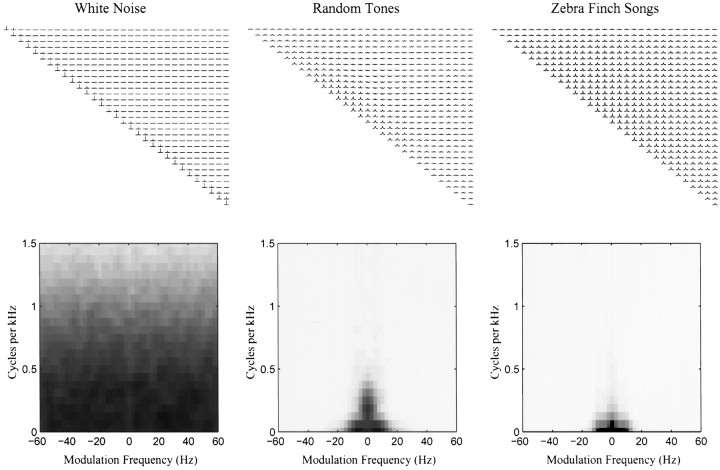
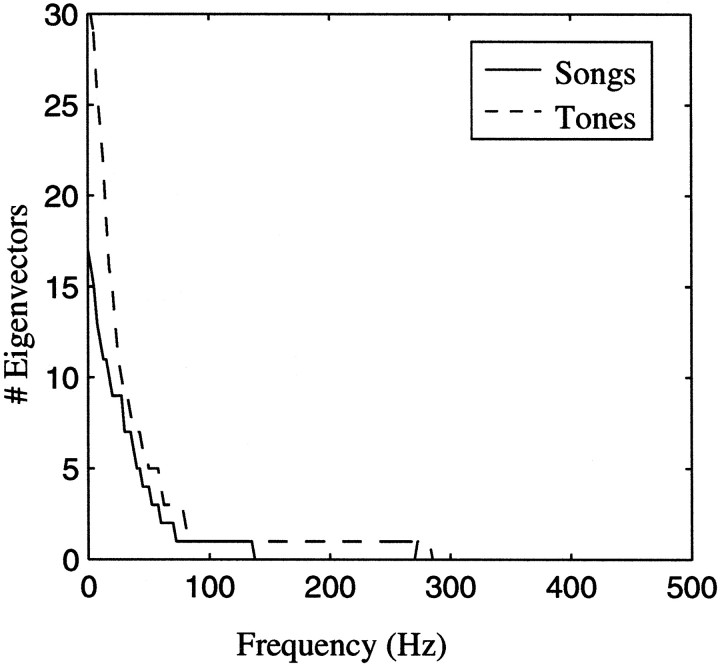
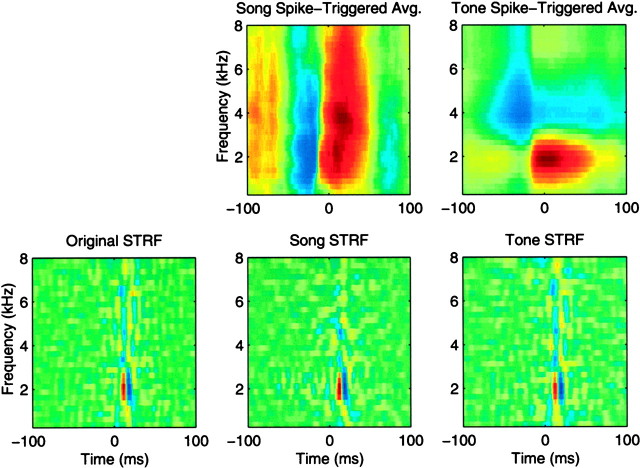
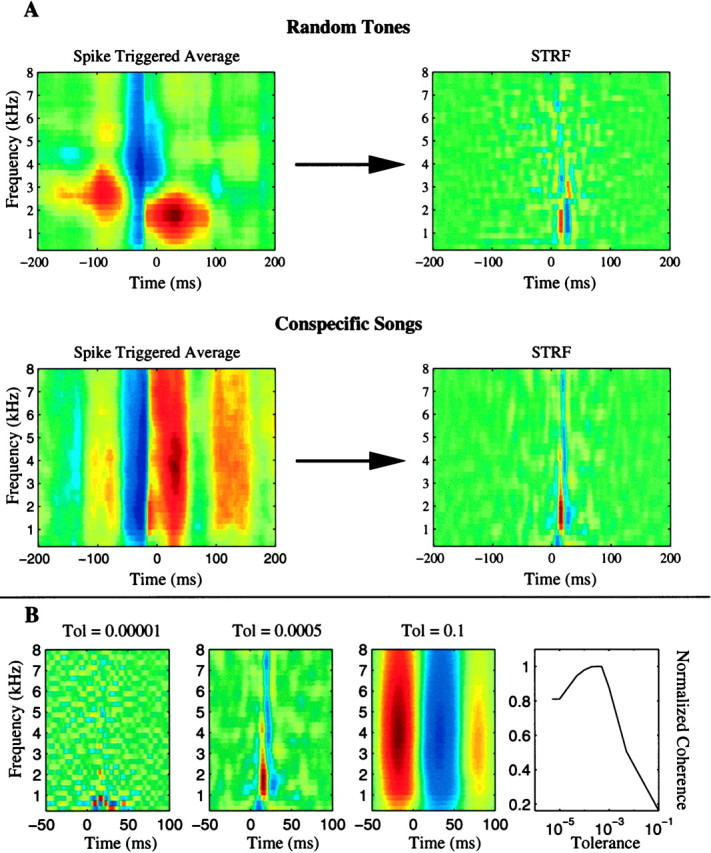
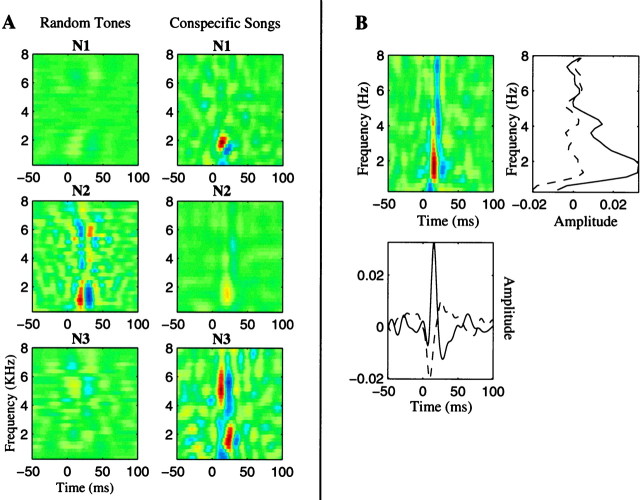
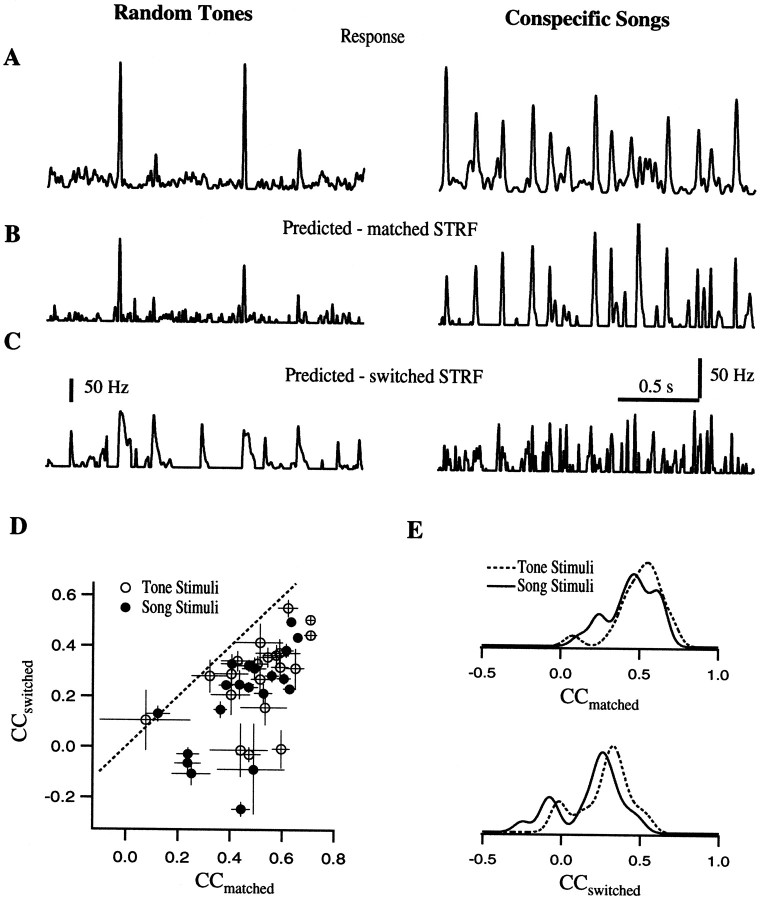
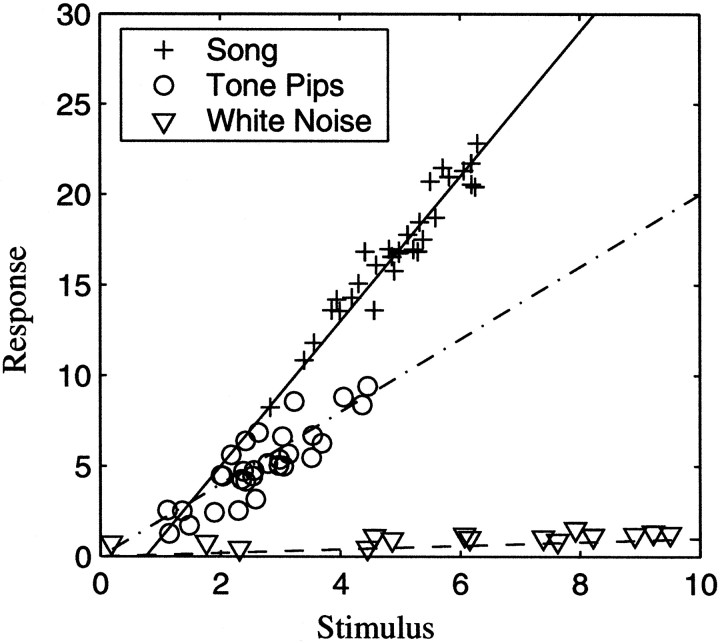
The stimulus-response function of many visual and auditory neurons has been described by a spatial-temporal receptive field (STRF), a linear model that for mathematical reasons has until recently been estimated with the reverse correlation method, using simple stimulus ensembles such as white noise. Such stimuli, however, often do not effectively activate high-level sensory neurons, which may be optimized to analyze natural sounds and images. We show that it is possible to overcome the simple-stimulus limitation and then use this approach to calculate the STRFs of avian auditory forebrain neurons from an ensemble of birdsongs. We find that in many cases the STRFs derived using natural sounds are strikingly different from the STRFs that we obtained using an ensemble of random tone pips. When we compare these two models by assessing their predictions of neural response to the actual data, we find that the STRFs obtained from natural sounds are superior. Our results show that the STRF model is an incomplete description of response properties of nonlinear auditory neurons, but that linear receptive fields are still useful models for understanding higher level sensory processing, as long as the STRFs are estimated from the responses to relevant complex stimuli.
Figures
References
-
- Aertsen AM, Johannesma PI. The spectro-temporal receptive field. A functional characteristic of auditory neurons. Biol Cybern. 1981a;42:133–143. - PubMed
-
- Aertsen AM, Johannesma PI. A comparison of the spectro-temporal sensitivity of auditory neurons to tonal and natural stimuli. Biol Cybern. 1981b;42:145–156. - PubMed
-
- Aertsen AM, Olders JH, Johannesma PI. Spectro-temporal receptive fields of auditory neurons in the grassfrog. III. Analysis of the stimulus-event relation for natural stimuli. Biol Cybern. 1981;39:195–209. - PubMed
-
- Boer ED, Kuyper P. Triggered correlation. IEEE Trans Biomed Eng. 1968;15:169–179. - PubMed
-
- Cai D, DeAngelis GC, Freeman RD. Spatiotemporal receptive field organization in the lateral geniculate nucleus of cats and kittens. J Neurophysiol. 1997;78:1045–1061. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources