

Afferent diversity and the organization of central vestibular pathways
- PMID: 10706428
- PMCID: PMC3731078
- DOI: 10.1007/s002210050033
Afferent diversity and the organization of central vestibular pathways
Abstract
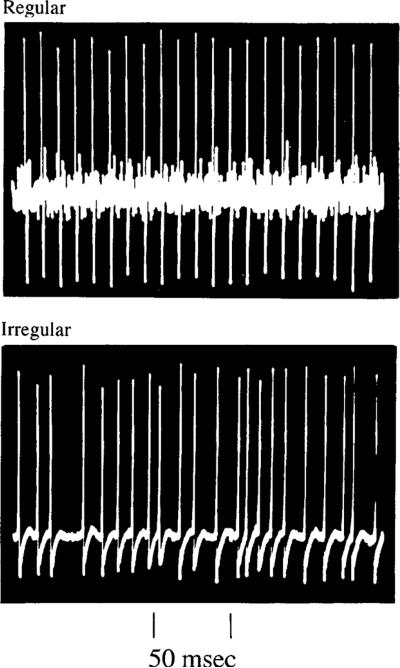
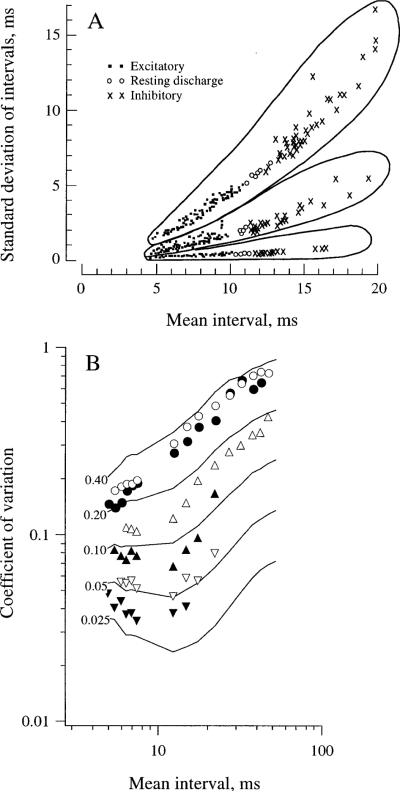
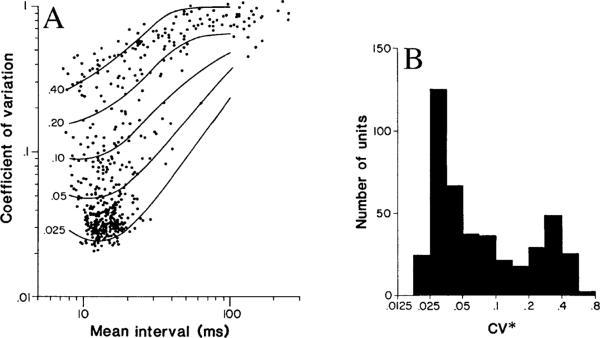
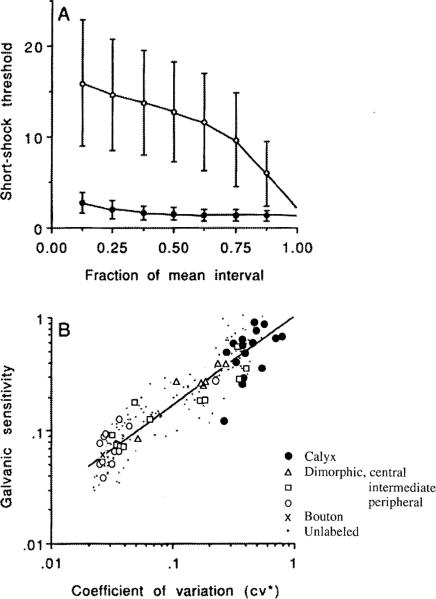
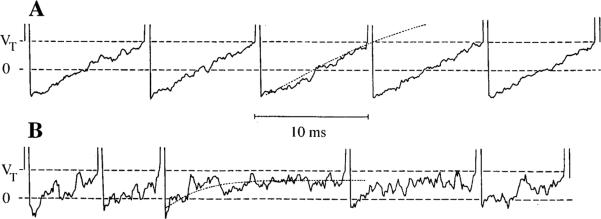
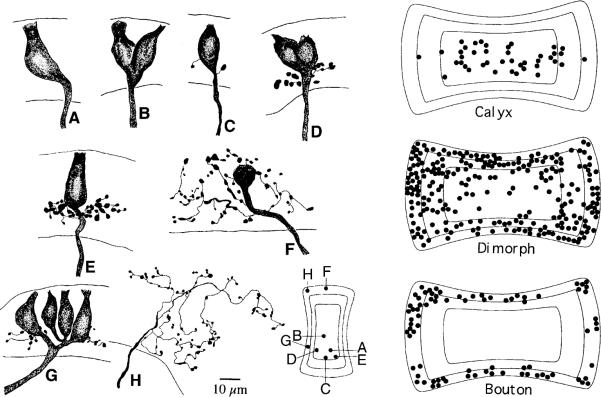
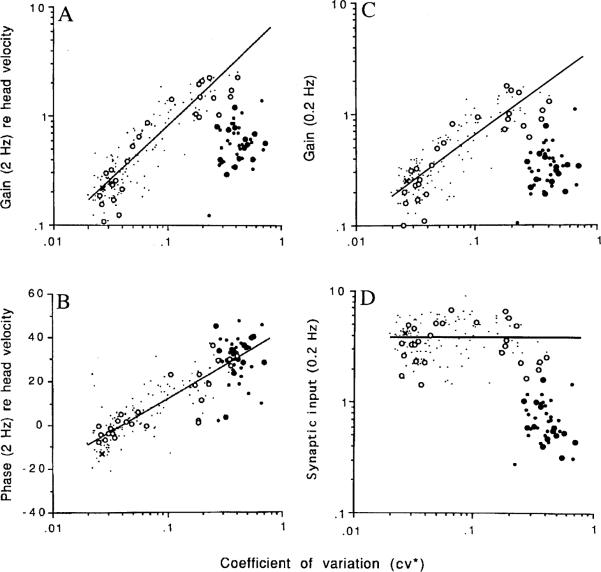
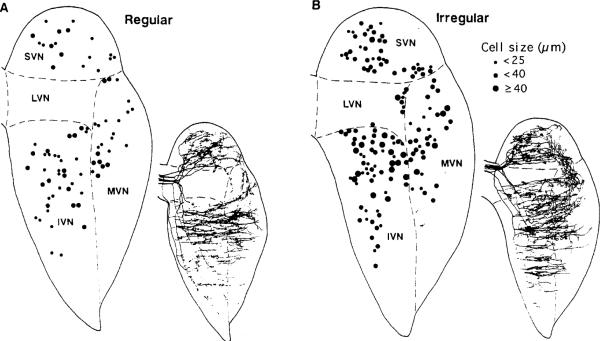
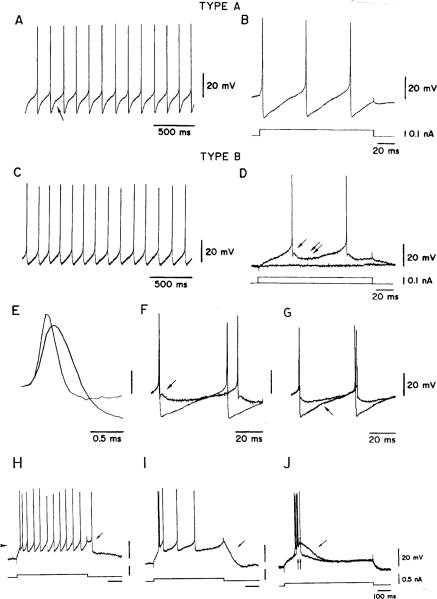
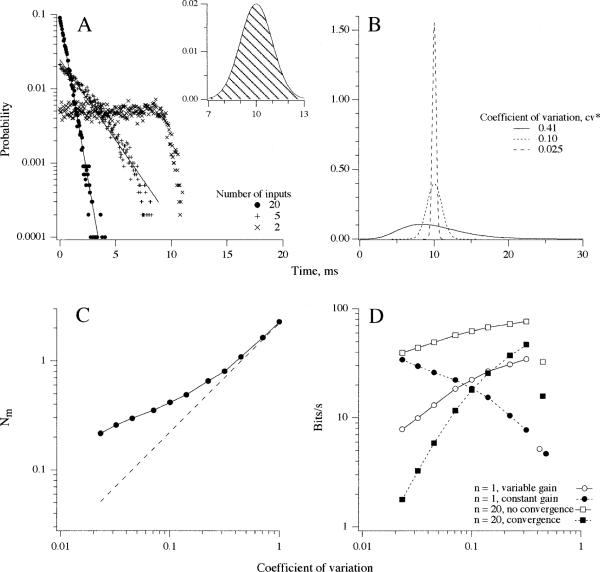
This review considers whether the vestibular system includes separate populations of sensory axons innervating individual organs and giving rise to distinct central pathways. There is a variability in the discharge properties of afferents supplying each organ. Discharge regularity provides a marker for this diversity since fibers which differ in this way also differ in many other properties. Postspike recovery of excitability determines the discharge regularity of an afferent and its sensitivity to depolarizing inputs. Sensitivity is small in regularly discharging afferents and large in irregularly discharging afferents. The enhanced sensitivity of irregular fibers explains their larger responses to sensory inputs, to efferent activation, and to externally applied galvanic currents, but not their distinctive response dynamics. Morphophysiological studies show that regular and irregular afferents innervate overlapping regions of the vestibular nuclei. Intracellular recordings of EPSPs reveal that some secondary vestibular neurons receive a restricted input from regular or irregular afferents, but that most such neurons receive a mixed input from both kinds of afferents. Anodal currents delivered to the labyrinth can result in a selective and reversible silencing of irregular afferents. Such a functional ablation can provide estimates of the relative contributions of regular and irregular inputs to a central neuron's discharge. From such estimates it is concluded that secondary neurons need not resemble their afferent inputs in discharge regularity or response dynamics. Several suggestions are made as to the potentially distinctive contributions made by regular and irregular afferents: (1) Reflecting their response dynamics, regular and irregular afferents could compensate for differences in the dynamic loads of various reflexes or of individual reflexes in different parts of their frequency range; (2) The gating of irregular inputs to secondary VOR neurons could modify the operation of reflexes under varying behavioral circumstances; (3) Two-dimensional sensitivity can arise from the convergence onto secondary neurons of otolith inputs differing in their directional properties and response dynamics; (4) Calyx afferents have relatively low gains when compared with irregular dimorphic afferents. This could serve to expand the stimulus range over which the response of calyx afferents remains linear, while at the same time preserving the other features peculiar to irregular afferents. Among those features are phasic response dynamics and large responses to efferent activation; (5) Because of the convergence of several afferents onto each secondary neuron, information transmission to the latter depends on the gain of individual afferents, but not on their discharge regularity.
Figures

References
-
- Aitkin LM, Irvine DRF, Webster WR. Central neural mechanisms of hearing. In: Darian-Smith I, editor. Handbook of physiology. Section I. The nervous system. Volume III. Sensory processes, Part 2. Williams & Wilkins; Baltimore: 1984. pp. 675–737.
-
- Angelaki DE, Perachio AA. Contribution of irregular semicircular canal afferents to the horizontal vestibuloocular response during constant velocity rotation. J Neurophysiol. 1993;69:996–999. - PubMed
-
- Angelaki DE, Perachio AA, Mustari MJ, Strunk CL. Role of irregular otolith afferents in the steady-state nystagmus during off-vertical axis rotation. J Neurophysiol. 1992;68:1895–1900. - PubMed
-
- Angelaki DE, McHenry MQ, Perachio AA, Dickman JD. Irregular otolith afferent inputs to the translational vestibuloocular reflex. Soc Neurosci Abstr. 1998;24:1411.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
