

A protein kinase activity associated with Epstein-Barr virus BGLF4 phosphorylates the viral early antigen EA-D in vitro
- PMID: 10708424
- PMCID: PMC111808
- DOI: 10.1128/jvi.74.7.3093-3104.2000
A protein kinase activity associated with Epstein-Barr virus BGLF4 phosphorylates the viral early antigen EA-D in vitro
Abstract
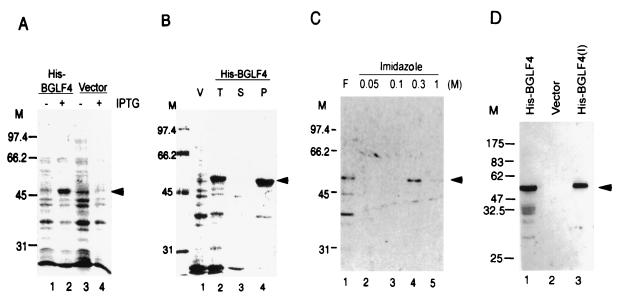
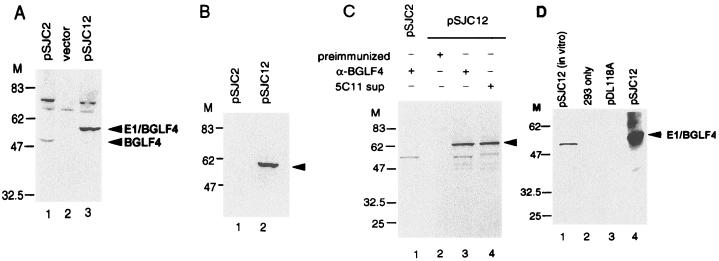
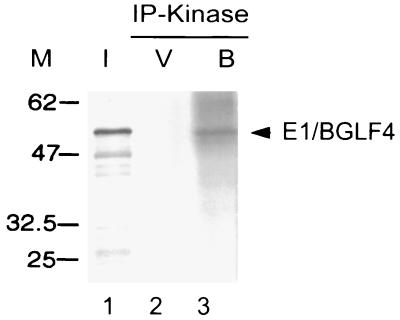
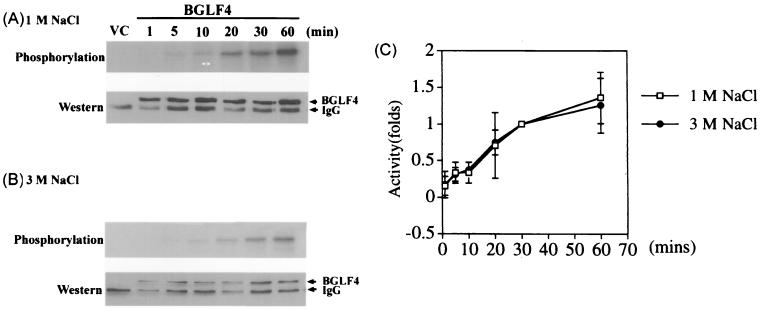
The Epstein-Barr virus (EBV) open reading frame BGLF4 was identified as a potential Ser/Thr protein kinase gene through the recognition of amino acid sequence motifs characteristic of conserved regions within the catalytic domains of protein kinases. In order to investigate this potential kinase activity, BGLF4 was expressed in Escherichia coli and the purified protein was used to generate a specific antiserum. Recombinant vaccinia virus vTF7-3, which expresses the T7 RNA polymerase, was used to infect 293 and 293T cells after transient transfection with a plasmid containing BGLF4 under the control of the T7 promoter. Autophosphorylation of the BGLF4 protein was demonstrated using the specific antiserum in an immune complex kinase assay. In addition, EBNA-1-tagged BGLF4 and EBNA-1 monoclonal antibody 5C11 were used to demonstrate the specificity of the kinase activity and to locate BGLF4 in the cytoplasm of transfected cells. Manganese ions were found to be essential for autophosphorylation of BGLF4, and magnesium can stimulate the activity. BGLF4 can utilize GTP, in addition to ATP, as a phosphate donor in this assay. BGLF4 can phosphorylate histone and casein in vitro. Among the potential viral protein substrates we examined, the EBV early antigen (EA-D, BMRF1), a DNA polymerase accessory factor and an important transactivator during lytic infection, was found to be phosphorylated by BGLF4 in vitro. Amino acids 1 to 26 of BGLF4, but not the predicted conserved catalytic domain, were found to be essential for autophosphorylation of BGLF4.
Figures




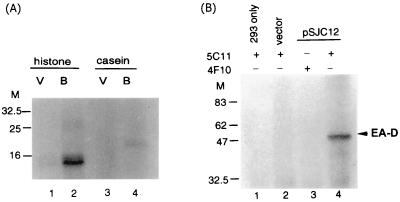
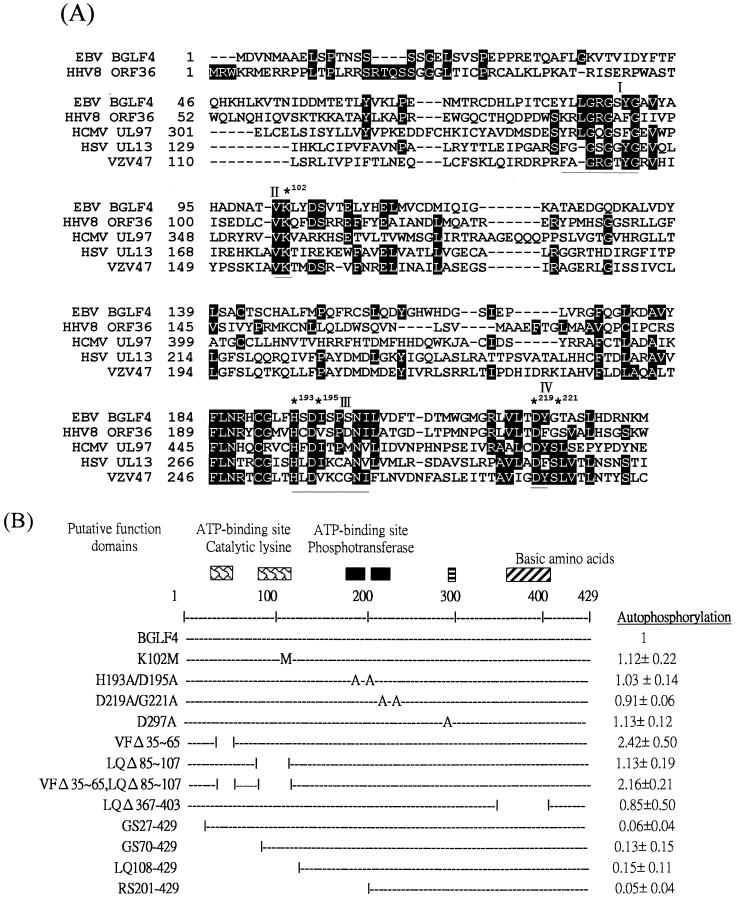


References
-
- Ali M A, Prakash S S, Jariwalla R. Localization of the antigenic sites and intrinsic protein kinase domain within a 300 amino acid segment of the ribonucleotide reductase large subunit from herpes simplex virus type 2. Virology. 1992;187:360–367. - PubMed
-
- Beaud G, Sharif A, Topa-Masse A, Leader D P. Ribosomal protein S2/Sa kinase purified from HeLa cells infected with vaccinia virus corresponds to the B1R protein kinase and phosphorylates in vitro the viral ssDNA-binding protein. J Gen Virol. 1994;75:283–293. - PubMed
-
- Branch D R, Mills G B. pp60c-src expression is induced by activation of normal human T lymphocytes. J Immunol. 1995;154:3678–3685. - PubMed
-
- Chee M S, Lawrence G L, Barrell B G. Alpha-, beta- beta- and gammaherpesviruses encode a putative phosphotransferase. J Gen Virol. 1989;70:1151–1160. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
