

A recombinant human parainfluenza virus type 3 (PIV3) in which the nucleocapsid N protein has been replaced by that of bovine PIV3 is attenuated in primates
- PMID: 10708435
- PMCID: PMC111819
- DOI: 10.1128/jvi.74.7.3188-3195.2000
A recombinant human parainfluenza virus type 3 (PIV3) in which the nucleocapsid N protein has been replaced by that of bovine PIV3 is attenuated in primates
Abstract
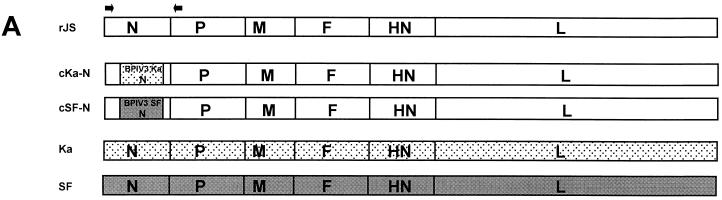
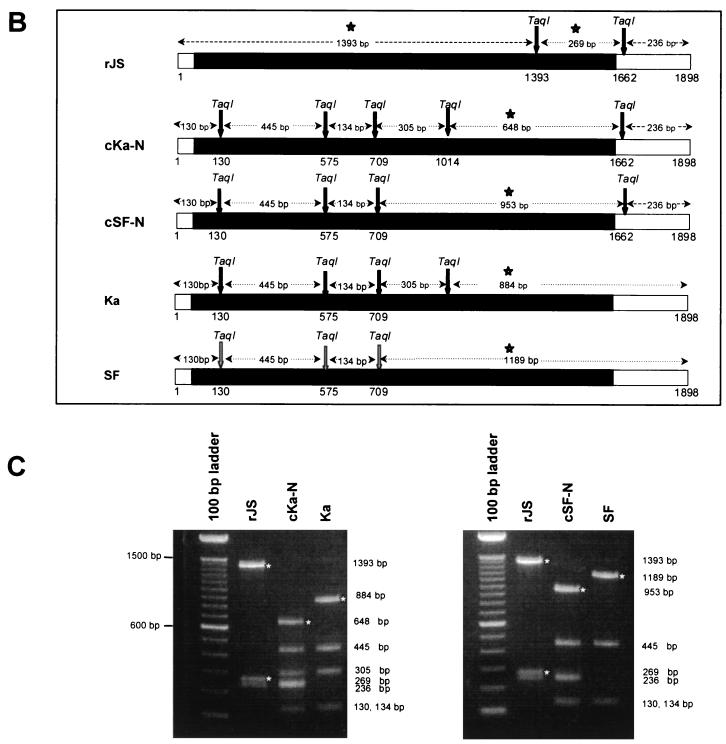
The shipping fever (SF) and Kansas (Ka) strains of bovine parainfluenza virus type 3 (BPIV3) are restricted in their replication in rhesus monkeys 100- to 1,000-fold compared to human parainfluenza virus type 3 (HPIV3), and the Ka strain also was shown to be attenuated in humans. To initiate an investigation of the genetic basis of the attenuation of BPIV3 in primates, we produced viable chimeric HPIV3 recombinants containing the nucleoprotein (N) open reading frame (ORF) from either BPIV3 Ka or SF in place of the HPIV3 N ORF. These chimeric recombinants were designated cKa-N and cSF-N, respectively. Remarkably, cKa-N and cSF-N grew to titers comparable to those of their HPIV3 and BPIV3 parents in LLC-MK2 monkey kidney and Madin-Darby bovine kidney cells. Thus, the heterologous nature of the N protein did not impede replication in vitro. However, cKa-N and cSF-N were each restricted in replication in rhesus monkeys to a similar extent as Ka and SF, respectively. This identified the BPIV3 N protein as a determinant of the host range restriction of BPIV3 in primates. These chimeras thus combine the antigenic determinants of HPIV3 with the host range restriction and attenuation phenotype of BPIV3. Despite their restricted replication in rhesus monkeys, the chimeric viruses induced a level of resistance to HPIV3 challenge in these animals which was indistinguishable from that conferred by immunization with HPIV3. The infectivity, attenuation, and immunogenicity of these BPIV3/HPIV3 chimeras suggest that the modified Jennerian approach described in the present report represents a novel method to design vaccines to protect against HPIV3-induced disease in humans.
Figures
References
-
- Bailly, J. E., J. M. McAuliffe, M. H. Skiadopoulos, P. L. Collins, and B. R. Murphy. Sequence determination and molecular analysis of two strains of bovine parainfluenza virus type 3 that are attenuated for primates. Virus Genes, in press. - PubMed
-
- Belshe R B, Hissom F K. Cold adaptation of parainfluenza virus type 3: induction of three phenotypic markers. J Med Virol. 1982;10:235–242. - PubMed
-
- Clements M L, Sears S D, Christina K, Murphy B R, Snyder M H. Comparison of the virologic and immunologic responses of volunteers to live avian-human influenza A H3N2 reassortant virus vaccines derived from two different avian influenza virus donors. J Clin Microbiol. 1989;27:219–222. - PMC - PubMed
-
- Clements M L, Subbarao E K, Fries L F, Karron R A, London W T, Murphy B R. Use of single-gene reassortant viruses to study the role of avian influenza A virus genes in attenuation of wild-type human influenza A virus for squirrel monkeys and adult human volunteers. J Clin Microbiol. 1992;30:655–662. - PMC - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
