

Recognition of a conserved class of RNA tetraloops by Saccharomyces cerevisiae RNase III
- PMID: 10716739
- PMCID: PMC16206
- DOI: 10.1073/pnas.97.7.3142
Recognition of a conserved class of RNA tetraloops by Saccharomyces cerevisiae RNase III
Abstract
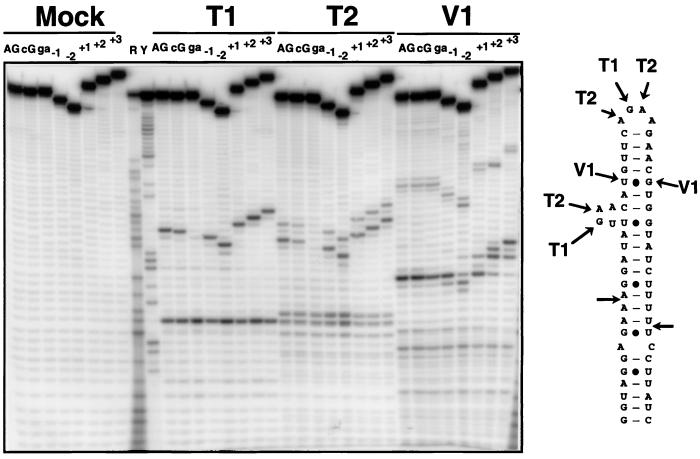
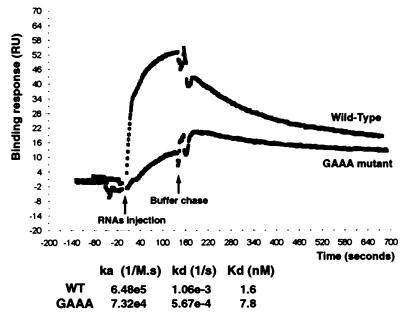
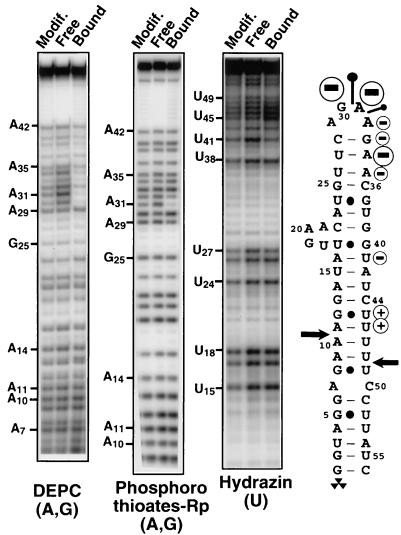
Ribonucleases III are double-stranded RNA (dsRNA) endonucleases required for the processing of a large number of prokaryotic and eukaryotic transcripts. Although the specificity of bacterial RNase III cleavage relies on antideterminants in the dsRNA, the molecular basis of eukaryotic RNase III specificity is unknown. All substrates of yeast RNase III (Rnt1p) are capped by terminal tetraloops showing the consensus AGNN and located within 13-16 bp to Rnt1p cleavage sites. We show that these tetraloops are essential for Rnt1p cleavage and that the distance to the tetraloop is the primary determinant of cleavage site selection. The presence of AGNN tetraloops also enhances Rnt1p binding, as shown by surface plasmon resonance monitoring and modification interference studies. These results define a paradigm of RNA loops and show that yeast RNase III behaves as a helical RNA ruler that recognizes these tetraloops and cleaves the dsRNA at a fixed distance to this RNA structure. These results also indicate that proteins belonging to the same class of RNA endonucleases require different structural elements for RNA cleavage.
Figures

References
-
- Robertson H D, Webster R E, Zinder N D. J Biol Chem. 1968;243:82–91. - PubMed
-
- Court D. In: Control of mRNA Stability. Brawerman G, Belasco J, editors. New York: Academic; 1993. pp. 70–116.
-
- Apirion D, Miczak A. BioEssays. 1993;15:113–120. - PubMed
-
- Nicholson A W. In: Ribonucleases: Structures and Functions. D'Alessio G, Riordan J F, editors. New York: Academic; 1997. pp. 1–49.
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
