

Role of gap junctions and EETs in endothelium-dependent hyperpolarization of porcine coronary artery
- PMID: 10725263
- PMCID: PMC1571957
- DOI: 10.1038/sj.bjp.0703188
Role of gap junctions and EETs in endothelium-dependent hyperpolarization of porcine coronary artery
Abstract
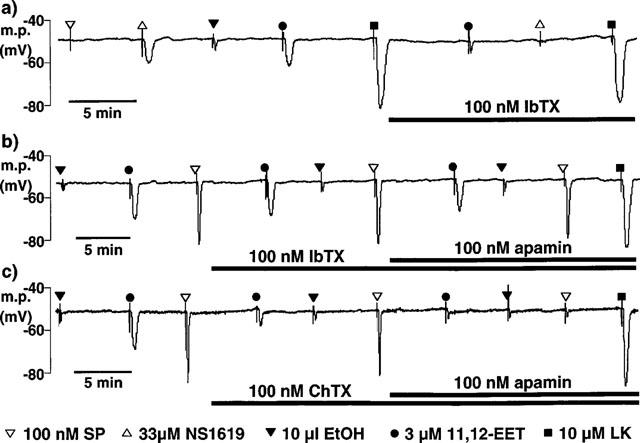
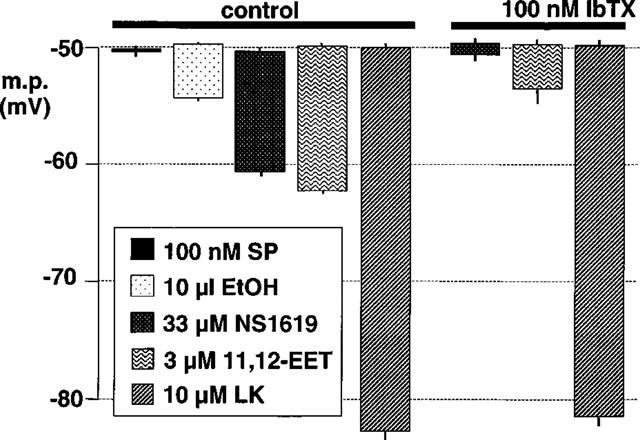
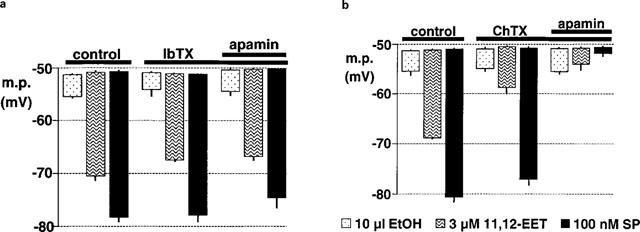
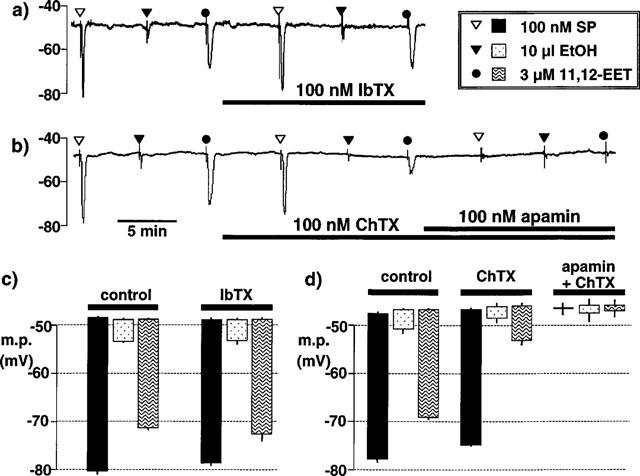
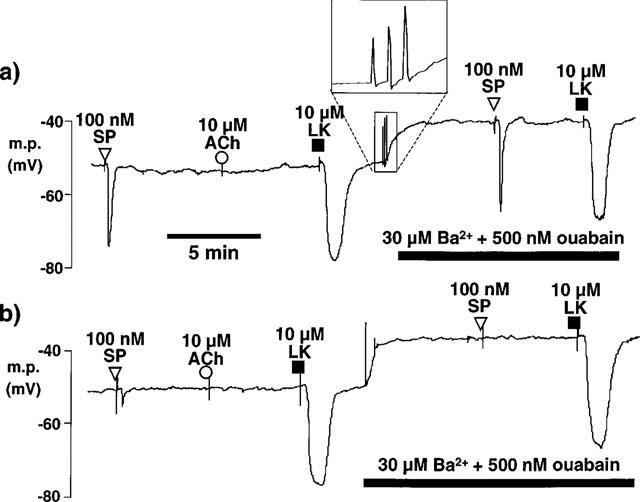
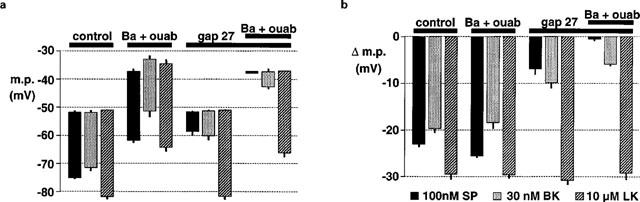
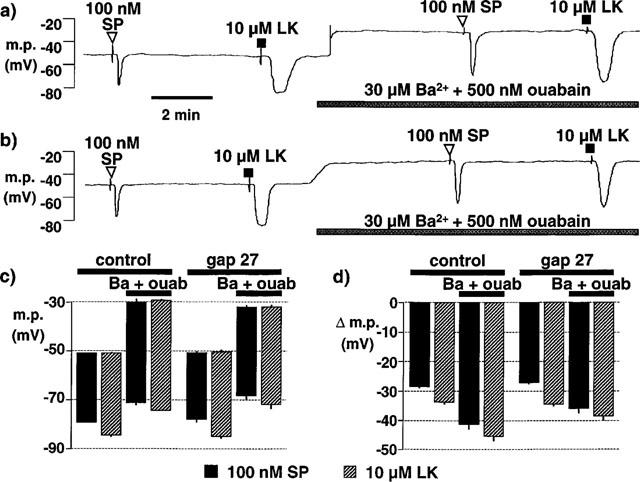
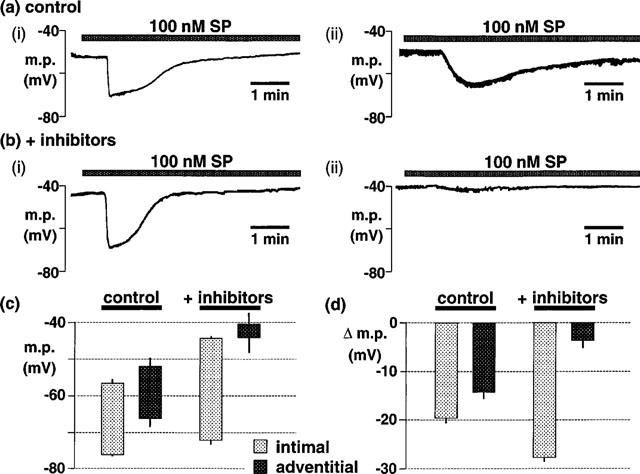
1. The effects of endothelium-derived hyperpolarizing factor (EDHF: elicited using substance P or bradykinin) were compared with those of 11,12-EET in pig coronary artery. Smooth muscle cells were usually impaled with microelectrodes through the adventitial surface. 2. Substance P (100 nM) and 11,12-EET (11,12-epoxyeicosatrienoic acid; 3 microM) hyperpolarized endothelial cells in intact arteries. These actions were unaffected by 100 nM iberiotoxin but were abolished by charybdotoxin plus apamin (each 100 nM). 3. Substance P (100 nM) and bradykinin (30 nM) hyperpolarized intact artery smooth muscle; Substance P had no effect after endothelium removal. 11,12-EET hyperpolarized de-endothelialized vessels by 12.6+/-0.3 mV, an effect abolished by 100 nM iberiotoxin. 4. 11,12-EET hyperpolarized intact arteries by 18.6+/-0.8 mV, an action reduced by iberiotoxin, which was ineffective against substance P. Hyperpolarizations to 11, 12-EET and substance P were partially inhibited by 100 nM charybdotoxin and abolished by further addition of 100 nM apamin. 5. 30 microM barium plus 500 nM ouabain depolarized intact artery smooth muscle but responses to substance P and bradykinin were unchanged. 500 microM gap 27 markedly reduced hyperpolarizations to substance P and bradykinin which were abolished in the additional presence of barium plus ouabain. 6. Substance P-induced hyperpolarizations of smooth muscle cells immediately below the internal elastic lamina were unaffected by gap 27, even in the presence of barium plus ouabain. 7. In pig coronary artery, 11,12-EET is not EDHF. Smooth muscle hyperpolarizations attributed to 'EDHF' are initiated by endothelial cell hyperpolarization involving charybdotoxin- (but not iberiotoxin) and apamin-sensitive K(+) channels. This may spread electrotonically via myoendothelial gap junctions but the involvement of an unknown endothelial factor cannot be excluded.
Figures
Similar articles
-
Bradykinin-induced, endothelium-dependent responses in porcine coronary arteries: involvement of potassium channel activation and epoxyeicosatrienoic acids.Br J Pharmacol. 2005 Jul;145(6):775-84. doi: 10.1038/sj.bjp.0706256. Br J Pharmacol. 2005. PMID: 15895105 Free PMC article.
-
Endothelium-dependent relaxation and hyperpolarization in guinea-pig coronary artery: role of epoxyeicosatrienoic acid.Br J Pharmacol. 1998 May;124(1):181-9. doi: 10.1038/sj.bjp.0701778. Br J Pharmacol. 1998. PMID: 9630358 Free PMC article.
-
Endothelium-independent, ouabain-sensitive relaxation of bovine coronary arteries by EETs.Am J Physiol Heart Circ Physiol. 2001 Mar;280(3):H1113-21. doi: 10.1152/ajpheart.2001.280.3.H1113. Am J Physiol Heart Circ Physiol. 2001. PMID: 11179054
-
Endothelial potassium channels, endothelium-dependent hyperpolarization and the regulation of vascular tone in health and disease.Clin Exp Pharmacol Physiol. 2004 Sep;31(9):641-9. doi: 10.1111/j.1440-1681.2004.04053.x. Clin Exp Pharmacol Physiol. 2004. PMID: 15479173 Review.
-
Endothelium-derived hyperpolarizing factor.Clin Exp Pharmacol Physiol. 1996 Dec;23(12):1082-90. doi: 10.1111/j.1440-1681.1996.tb01174.x. Clin Exp Pharmacol Physiol. 1996. PMID: 8977164 Review.
Cited by
-
Connexins and gap junctions in the EDHF phenomenon and conducted vasomotor responses.Pflugers Arch. 2010 May;459(6):897-914. doi: 10.1007/s00424-010-0830-4. Epub 2010 Apr 9. Pflugers Arch. 2010. PMID: 20379740 Review.
-
Dominant role of an endothelium-derived hyperpolarizing factor (EDHF)-like vasodilator in the ciliary vascular bed of the bovine isolated perfused eye.Br J Pharmacol. 2001 Oct;134(4):912-20. doi: 10.1038/sj.bjp.0704332. Br J Pharmacol. 2001. PMID: 11606333 Free PMC article.
-
Endothelium-dependent smooth muscle hyperpolarization: do gap junctions provide a unifying hypothesis?Br J Pharmacol. 2004 Mar;141(6):881-903. doi: 10.1038/sj.bjp.0705698. Br J Pharmacol. 2004. PMID: 15028638 Free PMC article. Review.
-
Characterization of a charybdotoxin-sensitive intermediate conductance Ca2+-activated K+ channel in porcine coronary endothelium: relevance to EDHF.Br J Pharmacol. 2002 Dec;137(8):1346-54. doi: 10.1038/sj.bjp.0705057. Br J Pharmacol. 2002. PMID: 12466245 Free PMC article.
-
Impaired vasodilation in the pathogenesis of hypertension: focus on nitric oxide, endothelial-derived hyperpolarizing factors, and prostaglandins.J Clin Hypertens (Greenwich). 2012 Apr;14(4):198-205. doi: 10.1111/j.1751-7176.2012.00606.x. J Clin Hypertens (Greenwich). 2012. PMID: 22458740 Free PMC article. Review.
References
-
- BÉNY J.-L., CONNAT J.L. An electron-microscope study of smooth muscle cell dye coupling in the pig coronary arteries. Role of gap junctions. Circ. Res. 1992;70:49–55. - PubMed
-
- BÉNY J.-L., GRIBI F. Dye and electrical coupling of endothelial cells in situ. Tissue Cell. 1989;21:797–802. - PubMed
-
- BÉNY J.-L., PACICCA C. Bidirectional electrical communication between smooth muscle and endothelial cells in the pig coronary artery. Am. J. Physiol. 1994;266:H1465–H1472. - PubMed
-
- CAMPBELL W.B., GEBREMEDHIN D., PRATT P.F., HARDER D.R. Identification of epoxyeicosatrienoic acids as endothelium-derived hyperpolarizing factors. Circ. Res. 1996;78:415–423. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
