

Essential function of Wnt-4 in mammary gland development downstream of progesterone signaling
Affiliations
- PMID: 10733525
- PMCID: PMC316462
Item in Clipboard
Essential function of Wnt-4 in mammary gland development downstream of progesterone signaling
Genes Dev.
.
Abstract
Female reproductive hormones control mammary gland morphogenesis. In the absence of the progesterone receptor (PR) from the mammary epithelium, ductal side-branching fails to occur. We can overcome this defect by ectopic expression of the protooncogene Wnt-1. Transplantation of mammary epithelia from Wnt-4(-)/(-) mice shows that Wnt-4 has an essential role in side-branching early in pregnancy. PR and Wnt-4 mRNAs colocalize to the luminal compartment of the ductal epithelium. Progesterone induces Wnt-4 in mammary epithelial cells and is required for increased Wnt-4 expression during pregnancy. Thus, Wnt signaling is essential in mediating progesterone function during mammary gland morphogenesis.
Figures
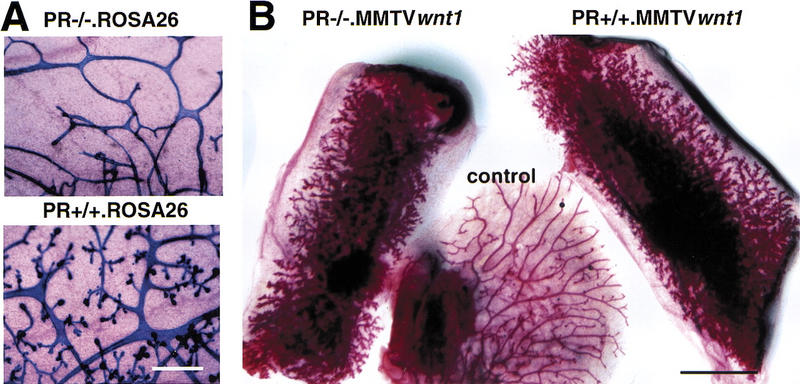
Side-branching in the presence of Wnt-1 and absence of the PR. (A) Intrinsic side-branching defect in PR−/− mammary epithelium. Mammary epithelium was harvested from PR−/− ROSA26 (top) and PR+/+ ROSA26 (bottom) 10-week-old female mice and engrafted to the cleared fat pads of 3-week-old F1 (129SV/C57B16) recipients. Shown are whole-mount preparations of mammary glands subjected to X-gal stain, from a recipient at day 12 of pregnancy (10 weeks after surgery). Results were similar to those reported previously. Bar, 400, μm. (B) Constitutive side-branching of MMTV Wnt-1tg irrespective of the PR status. Mammary epithelium was harvested from PR−/− MMTV Wnt-1tgand PR+/+ MMTV Wnt-1tb 10-week-old female mice and engrafted to the cleared fat pads of 3-week-old recipients. Shown are whole-mount preparations of mammary gland from a virgin RAG1−/− recipient 10 weeks after surgery. (Left) Inguinal fat pad engrafted with PR−/− MMTV Wnt-1tg mammary epithelium; (right) inguinal fat pad engrafted with PR+/+ MMTV Wnt-1tg mammary epithelium; (center) thoracic mammary gland, as an ungrafted endogenous control. Identical results were obtained in 16 independent grafts of PR−/− MMTV Wnt-1tg and control mammary epithelium. Bar, 5 mm.
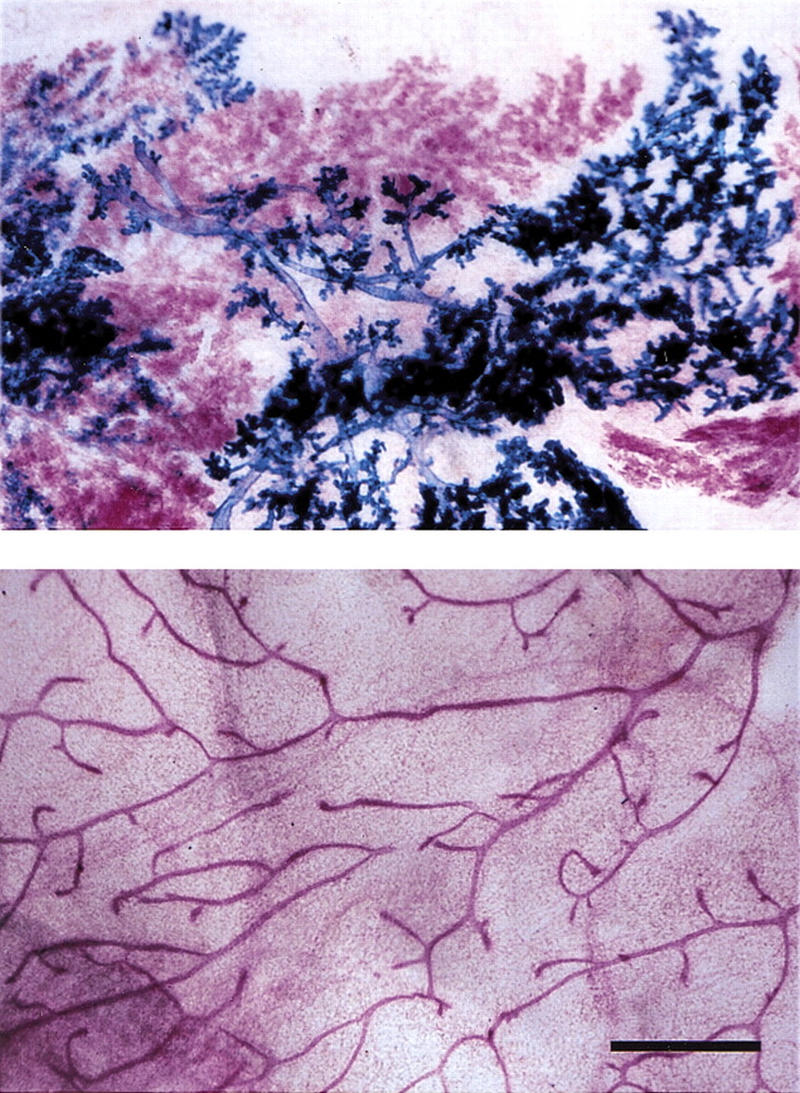
Paracrine induction of side-branching by Wnt-1. Primary mammary epithelial cells were derived from 10-week-old MMTV Wnt-1tg and ROSA26 females. After 5 days in vitro culture, the cells were trypsinized, mixed in a 1:1 ratio, and injected into cleared fat pads of 3-week-old RAG1−/− recipients. Mammary glands from RAG1−/− recipients at 10 weeks after surgery were subjected to X-gal stain, carmine alum counterstain, and mounted whole. (Top) Cleared fat pad reconstituted with a mixture of ROSA26 (blue) and MMTV Wnt1 (red) mammary epithelial cells; (bottom) ungrafted control, thoracic mammary gland. Bar, 500 μm. Wnt-1 overexpressing cells (red) induce premature side-branching in wild-type cells (blue).
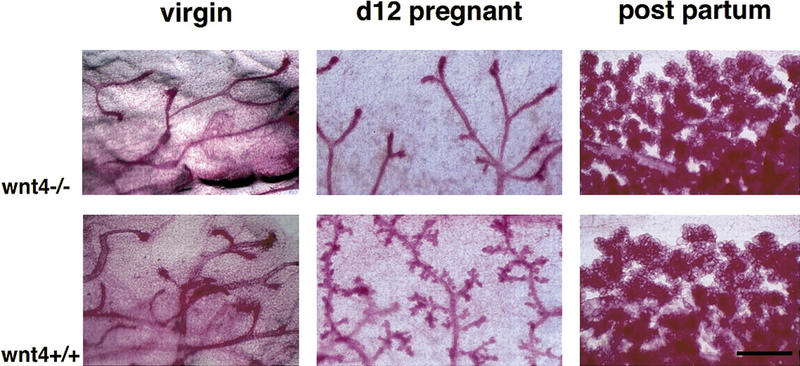
Function of Wnt-4 in the mammary epithelium at mid-pregnancy. Mammary buds were prepared from Wnt-4−/− and wild-type littermates (129SV/C57B16 mixed genetic background) at E14.5 and engrafted to the cleared fat pads of 3-week-old F1 (129SV/C57B16) recipients. At 10 weeks after surgery the mammary glands from virgin and impregnated recipients were analyzed by whole-mount preparation. (Top) Mutant grafts; (bottom) wild-type control grafts. Shown are mammary glands derived from a grafted virgin mouse (left), a mouse at day 12 of pregnancy (center), or at parturition (right). Wnt-4−/− epithelium fails to initiate side-branching at day 12 of pregnancy. Results were obtained with 4 virgin, 10 mid-pregnant, and 5 recipients at parturition. Bar 500 μm.
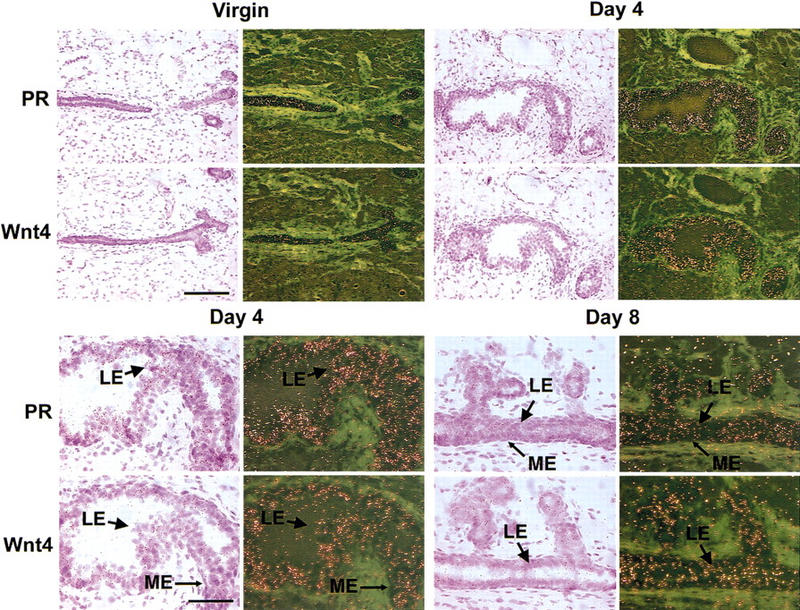
Coexpression of PR and Wnt-4 mRNAs in the luminal mammary epithelium. Mammary glands were harvested from a virgin adult female mouse and from mice at days 4 and 8 of pregnancy. The glands were processed for in situ hybridization; adjacent sections were hybridized with 35S-labeled antisense cRNA probes for PR or Wnt-4 and exposed for 7 days. Hematoxylin- and eosin-stained sections are shown next to the corresponding dark-field exposures. (Top) Coordinated induction of PR and Wnt-4 mRNAs during pregnancy. Low magnification (bar, 150 μm) of mammary gland showing increased signal intensity for both PR and Wnt-4 mRNAs in the ductal epithelium at 4 days of pregnancy vs. virgin. (Bottom) Colocalization of PR and Wnt-4 mRNA expression in the mammary luminal epithelium. High magnification (bar, 75 μm) of selections from mammary glands at days 4 and 8 of pregnancy showing that both PR and Wnt-4 mRNA expression localizes to the luminal epithelium (LE) and is absent from the myoepithelium (ME). Light green areas represent the dense fibrous stroma surrounding the mammary ducts.
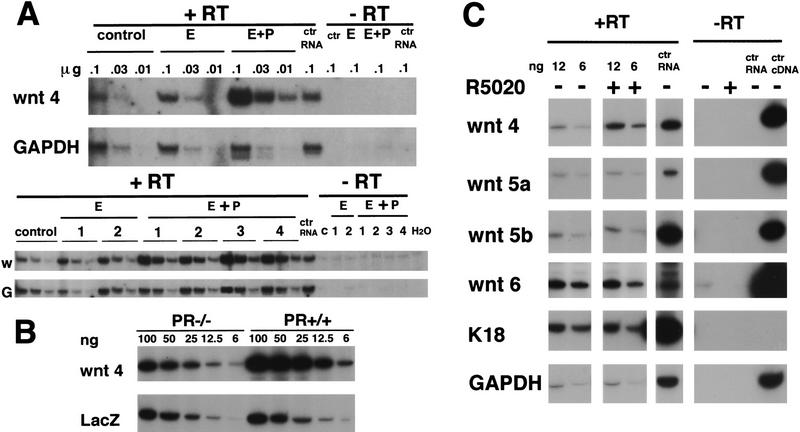
Induction of Wnt-4 expression in the mammary epithelium in vivo and in vitro by progesterone. (B) Quantification of Wnt-4 mRNA expression by semiquantitative PCR in mammary glands after 20 days of hormone treatment. Ten week-old virgin mice were ovarectomized. After 3 weeks they were injected for 20 days either with vehicle only (control), 10 μg of 17-β-estradiol (E)/day or 10 μg of estradiol and 100 μg of progesterone (E+P)/day. Total RNA was prepared from individual mammary glands, and samples in three serial dilutions, to ensure a linear signal response, were subjected to RT–PCR with primers specific for Wnt-4 or GAPDH. The same amounts of RNA in three serial dilutions were analyzed in each case. The undiluted RNA subjected to PCR amplification yielded no signal. Shown are two independent experiments, one comprising three mice (top) and one comprising seven mice (bottom). The products were quantified by densitometric scanning. The ratio of Wnt-4/GAPDH of the progesterone-treated samples was three- to fivefold higher than the 17-β-estradiol-treated samples. (B) Wnt-4 mRNA expression in mammary glands engrafted with PR−/− or PR+/+ mammary epithelium. Mammary epithelium was harvested from PR−/− ROSA26 and PR+/+ ROSA26 10-week-old female mice and engrafted to the cleared fat pads of 3-week-old F1 (129SV/C57B16) recipients. Six weeks after surgery the recipients were mated and the engrafted mammary glands were harvested at day 12 of pregnancy. RNA samples in five serial dilutions were subjected to RT–PCR with primers specific for Wnt-4 as in A. In parallel, RT–PCR was performed with lacZ-specific primers allowing normalization of the amount of transplanted epithelium. Densitometry reveals that the Wnt-4 signal is increased threefold in the PR−/− ROSA26 vs. the PR+/+ ROSA26 transplant. The same results were obtained in three independent experiments. (C) Wnt-4 mRNA expression in cultured primary mammary epithelial cells after progesterone exposure. Primary mammary epithelial cells were cultured on collagen-coated dishes for 3 days. RNA was harvested from untreated cells and cells after 8 hr of stimulation with the progesterone agonist R5020 (20 nm ) (P). Shown are two out of five serial dilutions of RNA subjected to RT–PCR with primers specific for Wnt-4, Wnt-5a, Wnt-6, keratin-18, and GAPDH. In each case, the undiluted RNA subjected to PCR ampliification without reverse transcription yielded no signal. Although the levels of Wnt-5a, Wnt-5b, Wnt-6, keratin-18, and GAPDH mRNA were unaffected by the treatment with R5020, the levels of Wnt-4 mRNA increased two- to threefold within 4–8 hours as confirmed in eight independent experiments.
References
-
- Bjornson C, Rietze R, Reynolds B, Magli M, Vescovi A. Turning brain into blood: A hematopoietic fate adopted by adult neural stem cells in vivo. Science. 1999;283:534–537. - PubMed
-
- Bocchinfuso W, Hively W, Couse J, Varmus H, Korach K. A mouse mammary tumor virus-Wnt-1 transgene induces mammary gland hyperplasia and tumorigenesis in mice lacking estrogen receptor-alpha. Cancer Res. 1999;59:1869–1876. - PubMed
-
- Bradbury JM, Edwards PA, Niemeyer CC, Dale TC. Wnt-4 expression induces a pregnancy-like growth pattern in reconstituted mammary glands in virgin mice. Dev Biol. 1995;170:553–563. - PubMed
-
- Cadigan K, Nusse R. Wnt signaling: A common theme in animal development. Genes & Dev. 1997;11:3286–3305. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials