

The KEN box: an APC recognition signal distinct from the D box targeted by Cdh1
- PMID: 10733526
- PMCID: PMC316466
The KEN box: an APC recognition signal distinct from the D box targeted by Cdh1
Abstract
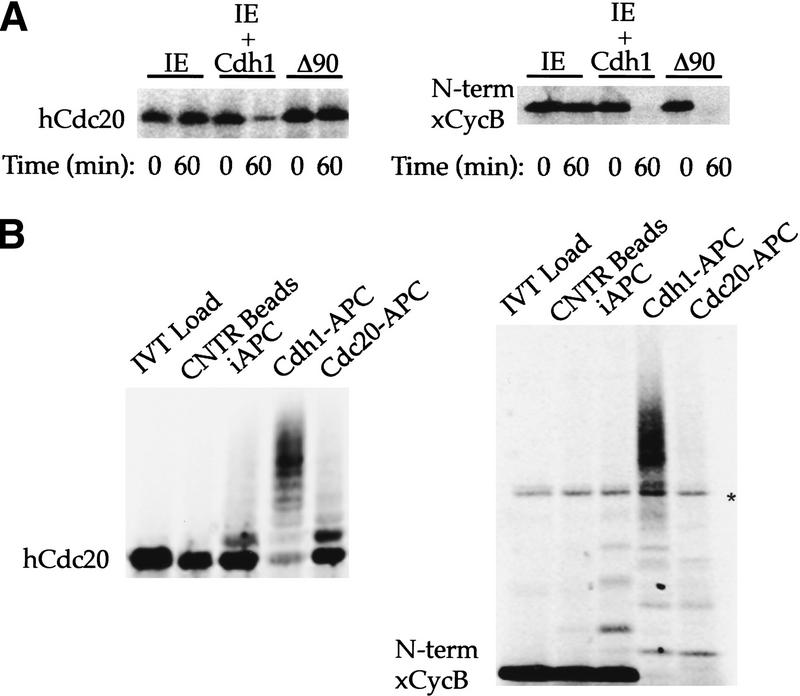
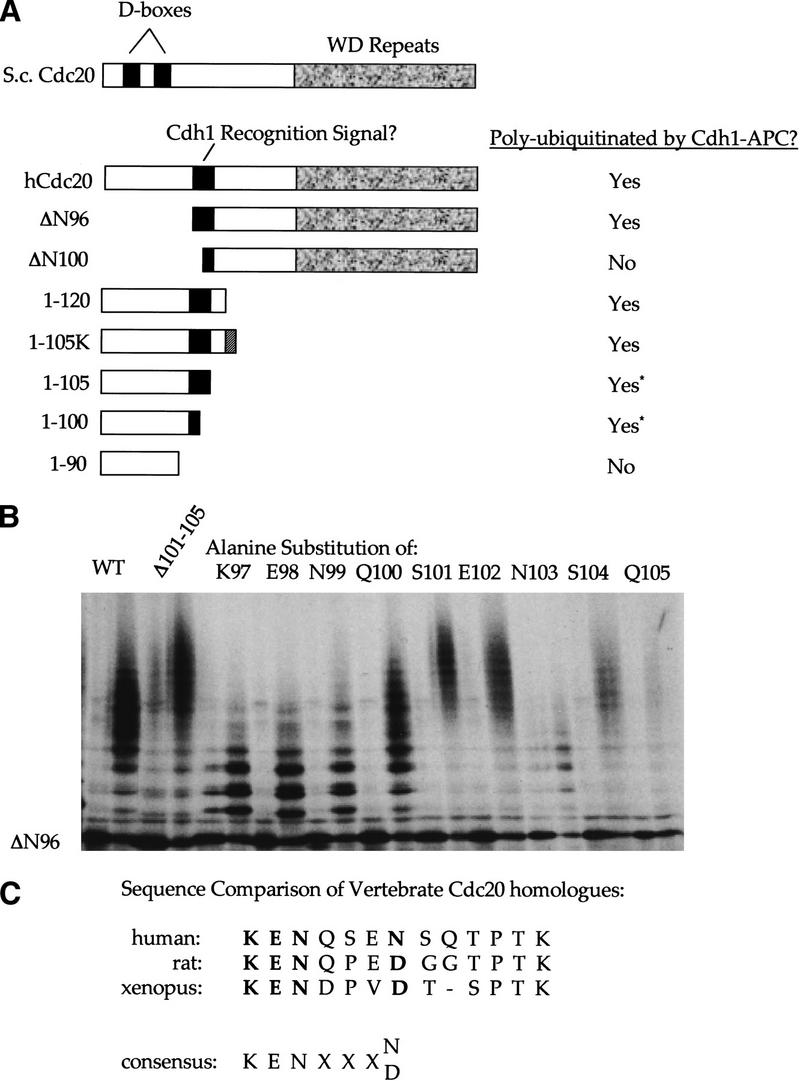
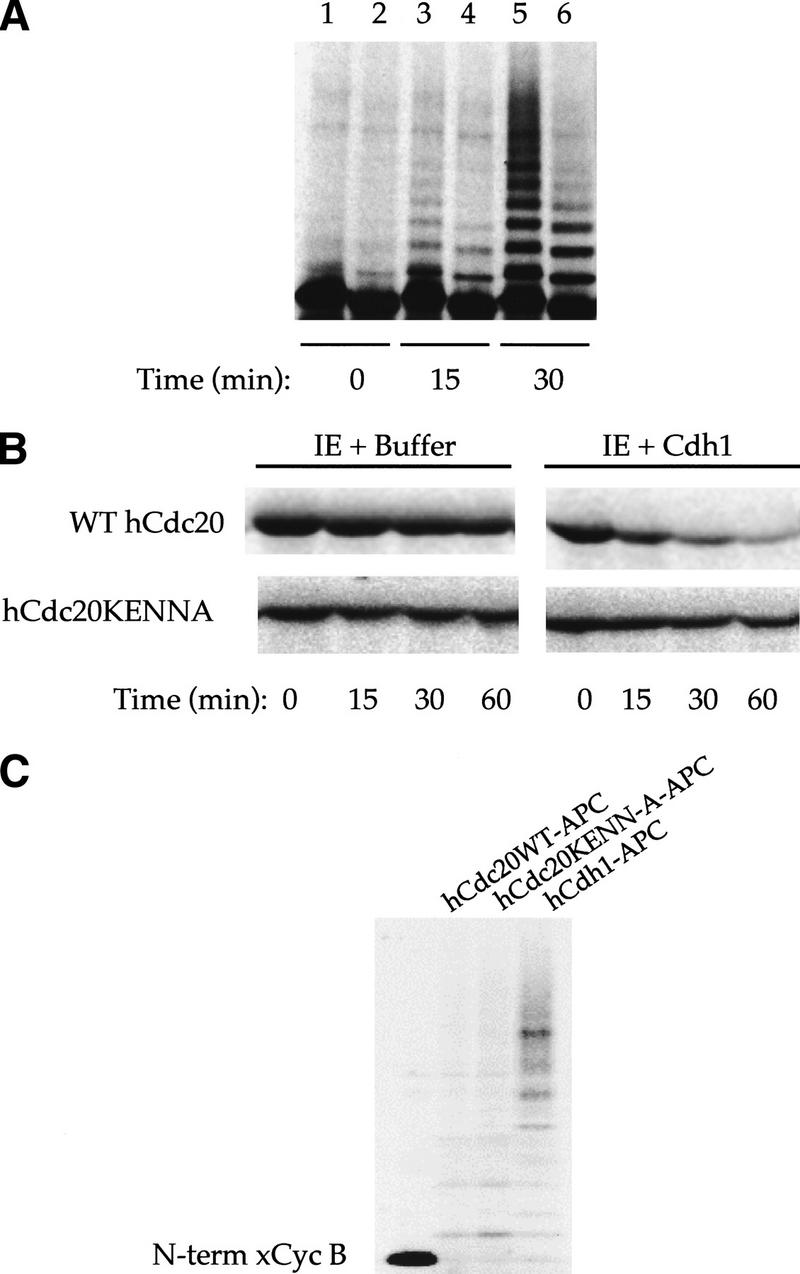
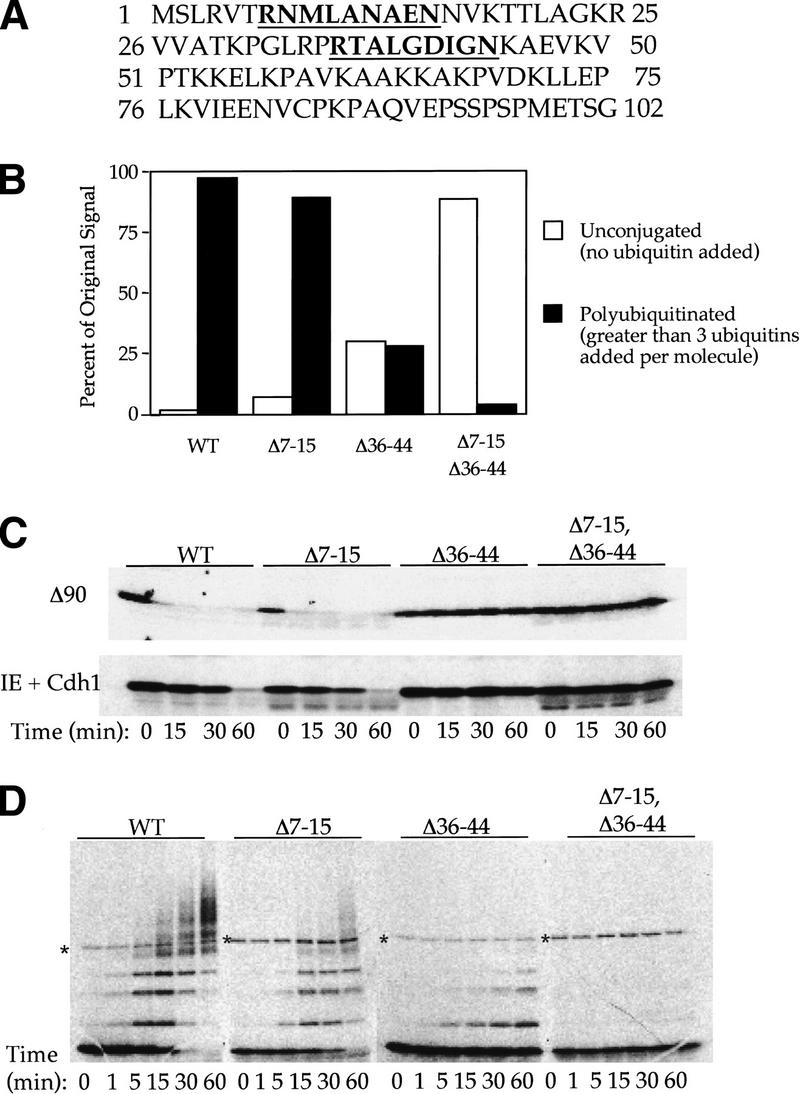
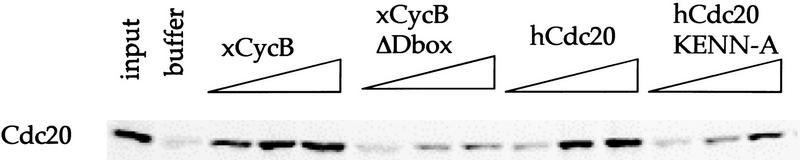
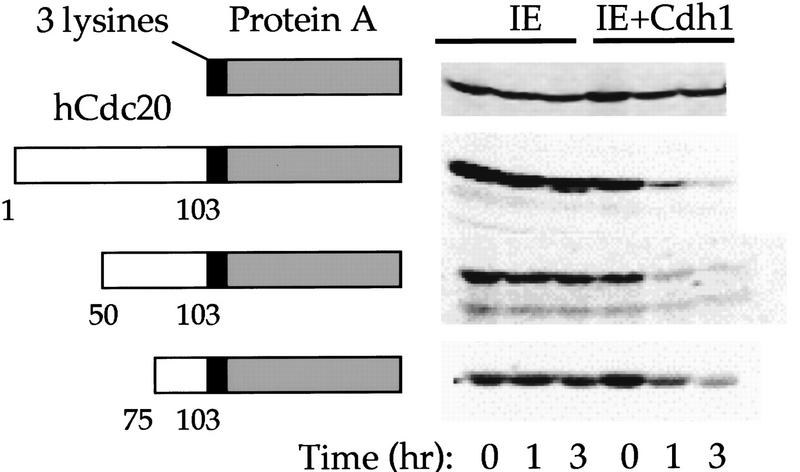
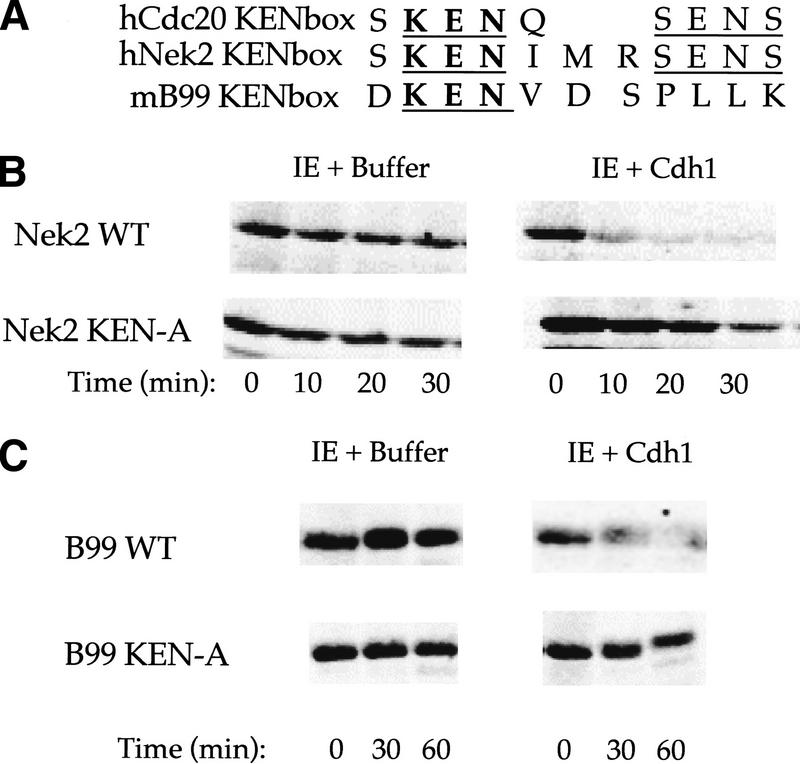
The ordered progression through the cell cycle depends on regulating the abundance of several proteins through ubiquitin-mediated proteolysis. Degradation is precisely timed and specific. One key component of the degradation system, the anaphase promoting complex (APC), is a ubiquitin protein ligase. It is activated both during mitosis and late in mitosis/G(1), by the WD repeat proteins Cdc20 and Cdh1, respectively. These activators target distinct sets of substrates. Cdc20-APC requires a well-defined destruction box (D box), whereas Cdh1-APC confers a different and as yet unidentified specificity. We have determined the sequence specificity for Cdh1-APC using two assays, ubiquitination in a completely defined and purified system and degradation promoted by Cdh1-APC in Xenopus extracts. Cdc20 is itself a Cdh1-APC substrate. Vertebrate Cdc20 lacks a D box and therefore is recognized by Cdh1-APC through a different sequence. By analysis of Cdc20 as a substrate, we have identified a new recognition signal. This signal, composed of K-E-N, serves as a general targeting signal for Cdh1-APC. Like the D box, it is transposable to other proteins. Using the KEN box as a template, we have identified cell cycle genes Nek2 and B99 as additional Cdh1-APC substrates. Mutation in the KEN box stabilizes all three proteins against ubiquitination and degradation.
Figures
References
-
- Bischoff J, Plowman GD. The Auror/Ip1p kinase family: Regulators of chromosome segregation and cytokinesis. Trends Cell Biol. 1999;9:454–459. - PubMed
-
- Ciechanover A. The ubiquitin-proteasome proteolytic pathway. Cell. 1994;79:13–21. - PubMed
-
- Cohen-Fix O, Peters J-M, Kirschner MW, Koshland D. Anaphase initiation in Saccharomyces cerevisiae is controlled by the APC-dependent degradation of the anaphase inhibitor Pds1p. Genes & Dev. 1996;10:3081–3093. - PubMed
-
- Coux O, Tanaka K, Goldberg AL. Structure and functions of the 20S and 26S proteasomes. Annu Rev Biochem. 1996;65:801–847. - PubMed
-
- Fang G, Yu H, Kirschner MW. Direct binding of CDC20 protein family members activates the anaphase-promoting complex in Mitosis and G1. Mol Cell. 1998;2:163–171. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous