

Multiple ras downstream pathways mediate functional repression of the homeobox gene product TTF-1
- PMID: 10733581
- PMCID: PMC85494
- DOI: 10.1128/MCB.20.8.2783-2793.2000
Multiple ras downstream pathways mediate functional repression of the homeobox gene product TTF-1
Abstract
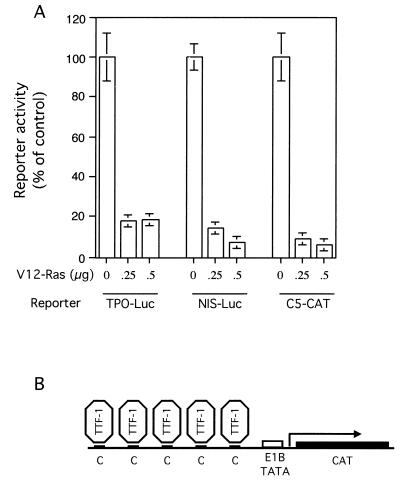
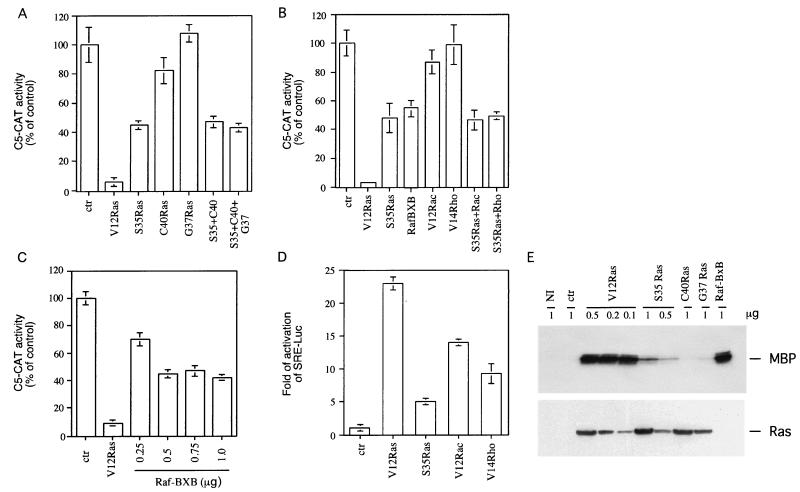
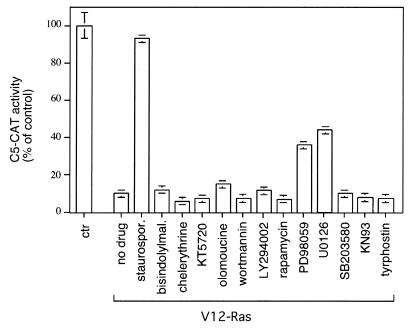
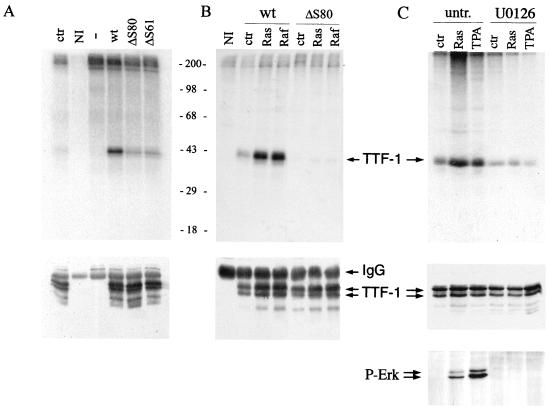
Expression of oncogenic Ras in thyroid cells results in loss of expression of several thyroid-specific genes and inactivation of TTF-1, a homeodomain-containing transcription factor required for normal development of the thyroid gland. In an effort to understand how signal transduction pathways downstream of Ras may be involved in suppression of the differentiated phenotype, we have tested mutants of the Ras effector region for their ability to affect TTF-1 transcriptional activity in a transient-transfection assay. We find that V12S35 Ras, a mutant known to interact specifically with Raf but not with RalGDS or phosphatidylinositol 3-kinase (PI3 kinase) inhibits TTF-1 activity. Expression of an activated form of Raf (Raf-BXB) also inhibits TTF-1 function to a similar extent, while the MEK inhibitors U0126 and PD98059 partially relieve Ras-mediated inactivation of TTF-1, suggesting that the extracellular signal-regulated kinase (ERK) pathway is involved in this process. Indeed, ERK directly phosphorylates TTF-1 at three serine residues, and concomitant mutation of these serines to alanines completely abolishes ERK-mediated phosphorylation both in vitro and in vivo. Since activation of the Raf/MEK/ERK pathway accounts for only part of the activity elicited by oncogenic Ras on TTF-1, other downstream pathways are likely to be involved in this process. We find that activation of PI3 kinase, Rho, Rac, and RalGDS has no effect on TTF-1 transcriptional activity. However, a poorly characterized Ras mutant, V12N38 Ras, can partially repress TTF-1 transcriptional activity through an ERK-independent pathway. Importantly, concomitant expression of constitutive activated Raf and V12N38 Ras results in almost complete loss of TTF-1 activity. Our data indicate that the Raf/MEK/ERK cascade may act in concert with an as-yet-uncharacterized signaling pathway activated by V12N38 Ras to repress TTF-1 function and ultimately to inhibit thyroid cell differentiation.
Figures
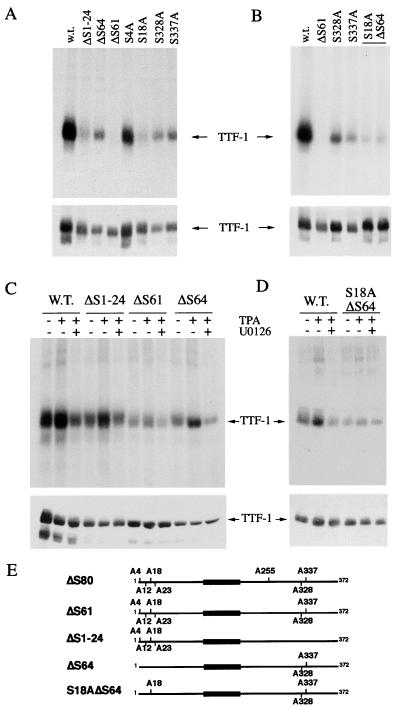
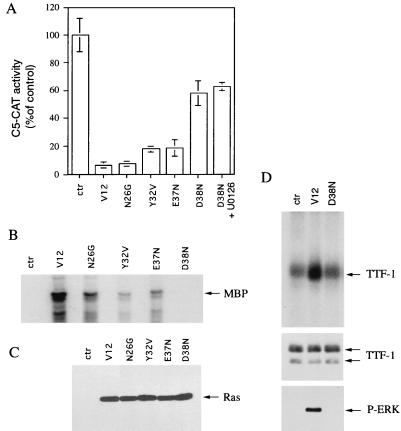
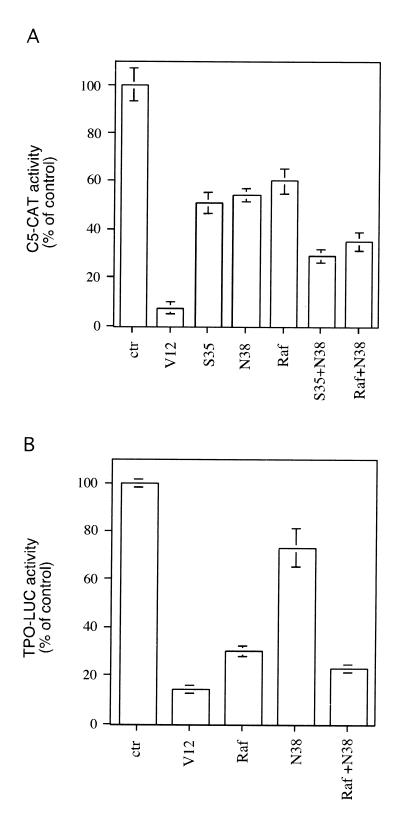
References
-
- Akasaka K, Tamada M, Wang F, Kariya K, Shima F, Kikuchi A, Yamamoto M, Shirouzu M, Yokoyama S, Kataoka T. Differential structural requirements for interaction of Ras protein with its distinct downstream effectors. J Biol Chem. 1996;271:5353–5360. - PubMed
-
- Baker S J, Markowitz S, Fearon E R, Willson J K, Vogelstein B. Suppression of human colorectal carcinoma cell growth by wild-type p53. Science. 1990;249:912–915. - PubMed
-
- Bos J L. The ras gene family and human carcinogenesis. Mutat Res. 1988;195:255–271. - PubMed
-
- Brasier A R, Tate J E, Habener J F. Optimized use of the firefly luciferase assay as a reporter gene in mammalian cell lines. Biotechniques. 1989;7:1116–1122. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous