

Dpb11 controls the association between DNA polymerases alpha and epsilon and the autonomously replicating sequence region of budding yeast
- PMID: 10733584
- PMCID: PMC85497
- DOI: 10.1128/MCB.20.8.2809-2817.2000
Dpb11 controls the association between DNA polymerases alpha and epsilon and the autonomously replicating sequence region of budding yeast
Abstract
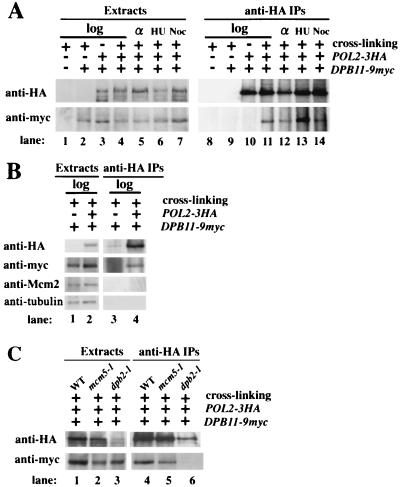
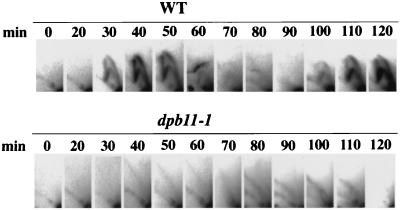
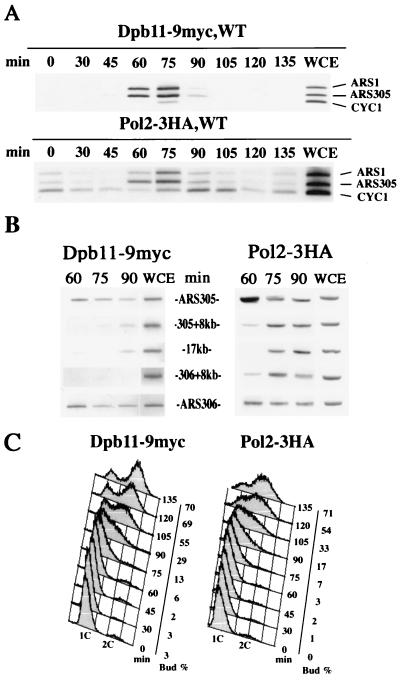
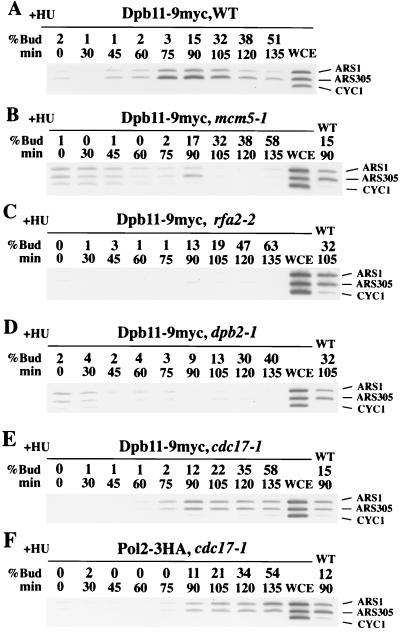
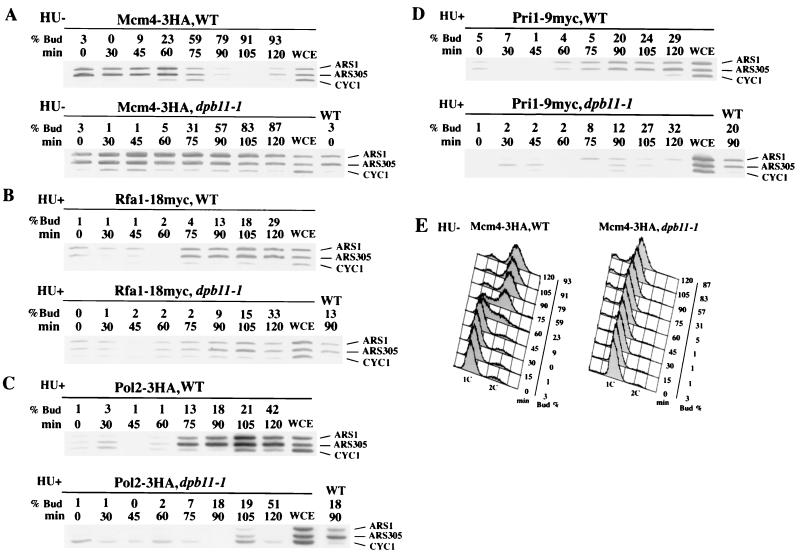
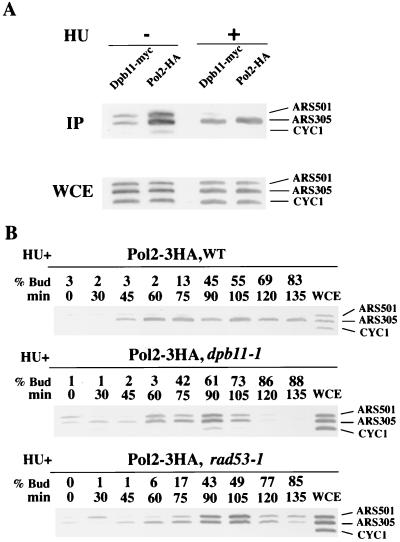
Dpb11 is required for chromosomal DNA replication and the S-phase checkpoint in Saccharomyces cerevisiae. Here, we report detection of a physical complex containing Dpb11 and DNA polymerase epsilon (Dpb11-Polepsilon complex). During the S phase of the cell cycle, Dpb11 associated preferentially with DNA fragments containing autonomously replicating sequences (ARSs), at the same time as Polepsilon associated with these fragments. Association of Dpb11 and Polepsilon with these fragments was mutually dependent, suggesting that the Dpb11-Polepsilon complex associates with the ARS. Moreover, Dpb11 was required for the association of Polalpha-primase with the fragments. Thus, it seems likely that association of the Dpb11-Polepsilon complex with the ARS fragments is required for the association of the Polalpha-primase complex. Hydroxyurea inhibits late-origin firing in S. cerevisiae, and the checkpoint genes, RAD53 and MEC1, are involved in this inhibition. In the presence of hydroxyurea at temperatures permissive for cell growth, Polepsilon in dpb11-1 cells associated with early- and late-origin fragments. In wild-type cells, however, it associated only with early-origin fragments. This indicates that Dpb11 may also be involved in the regulation of late-origin firing. Overall, these results suggest that Dpb11 controls the association between DNA polymerases alpha and epsilon and the ARS.
Figures
References
-
- Aparicio O M, Weinstein D M, Bell S P. Components and dynamics of DNA replication complexes in S. cerevisiae: redistribution of MCM proteins and Cdc45p during S phase. Cell. 1997;91:59–69. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases