

Production of pectate lyases and cellulases by Chryseomonas luteola strain MFCL0 depends on the growth temperature and the nature of the culture medium: evidence for two critical temperatures
- PMID: 10742239
- PMCID: PMC92020
- DOI: 10.1128/AEM.66.4.1538-1543.2000
Production of pectate lyases and cellulases by Chryseomonas luteola strain MFCL0 depends on the growth temperature and the nature of the culture medium: evidence for two critical temperatures
Abstract
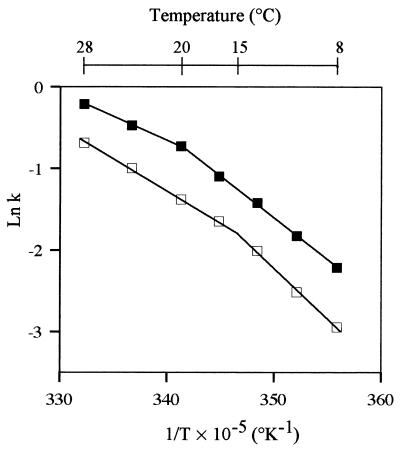
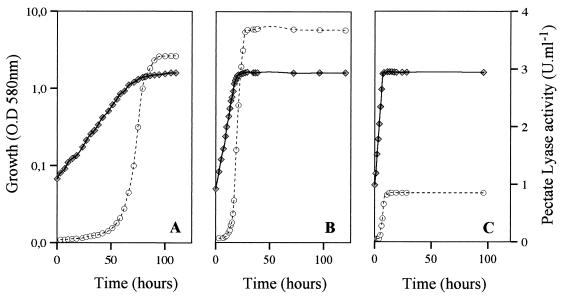
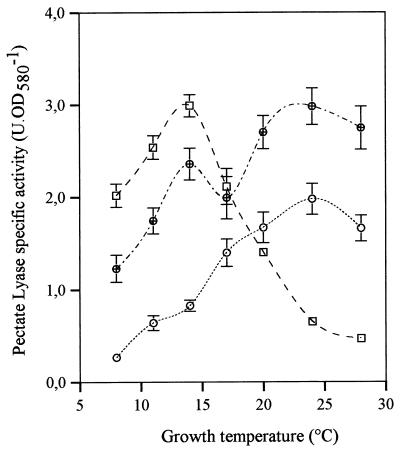
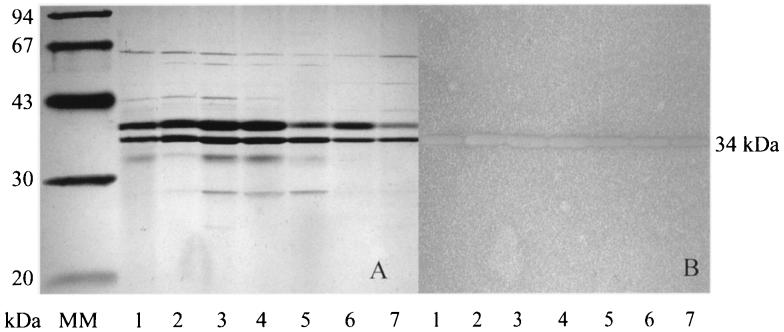
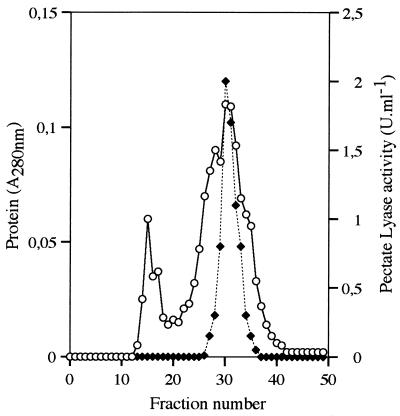
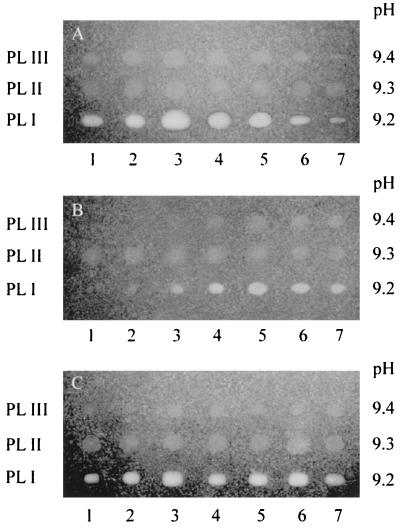
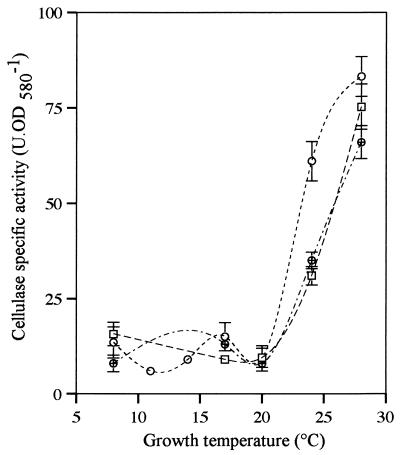
Several extracellular enzymes that are responsible for plant tissue maceration were detected in culture supernatant of the psychrotrophic bacterium Chryseomonas luteola MFCL0. Isoelectrofocusing experiments showed that pectate lyase (PL) activity resulted from the cumulative action of three major isoenzymes, designated PLI, PLII, and PLIII. Cellulolytic activity was also detected in culture supernatants. These enzymes exhibited different behaviors with respect to growth temperature. PLII was not regulated by temperature, whereas PLI and PLIII were regulated similarly by growth temperature. Maximal levels of PLI and PLIII were produced at 14 degrees C when cells were grown in polygalacturonate-containing synthetic medium and at around 20 to 24 degrees C in nutrient broth. In contrast, thermoregulation of cellulolytic activity production differed from thermoregulation of PL. The level of cellulolytic activity was low in all media at temperatures up to 20 degrees C, and then it increased dramatically until the temperature was 28 degrees C, which is the optimal temperature for growth of C. luteola. Previously, we defined the critical temperature by using the modified Arrhenius equation to characterize bacterial behavior. This approach consists of monitoring changes in the maximal specific growth rate as a function of temperature. Our most striking result was the finding that the temperature at which maximum levels of PLI and PLIII were produced in two different media was the same as the critical temperature for growth observed in these two media.
Figures
References
-
- Barras F, van Gijsegem F, Chatterjee A K. Extracellular enzymes and pathogenesis of soft-rot Erwinia. Annu Rev Phytopathol. 1994;32:201–234.
-
- Beguin P, Aubert J P. The biological degradation of cellulose. FEMS Microbiol Rev. 1994;13:25–58. - PubMed
-
- Bertheau Y, Madgidi-Hervan E, Kotoujanski A, Nguyen-The C, Andro T, Coleno A. Detection of depolymerase isoenzymes after electrophoresis or electrofocusing, or in titration curves. Anal Biochem. 1984;139:383–389. - PubMed
-
- Brocklehurst T F, Lund B M. Properties of pseudomonads causing spoilage of vegetables stored at low temperature. J Appl Bacteriol. 1981;50:259–266.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
