

Synaptic control of motoneuronal excitability
- PMID: 10747207
- PMCID: PMC4764886
- DOI: 10.1152/physrev.2000.80.2.767
Synaptic control of motoneuronal excitability
Abstract
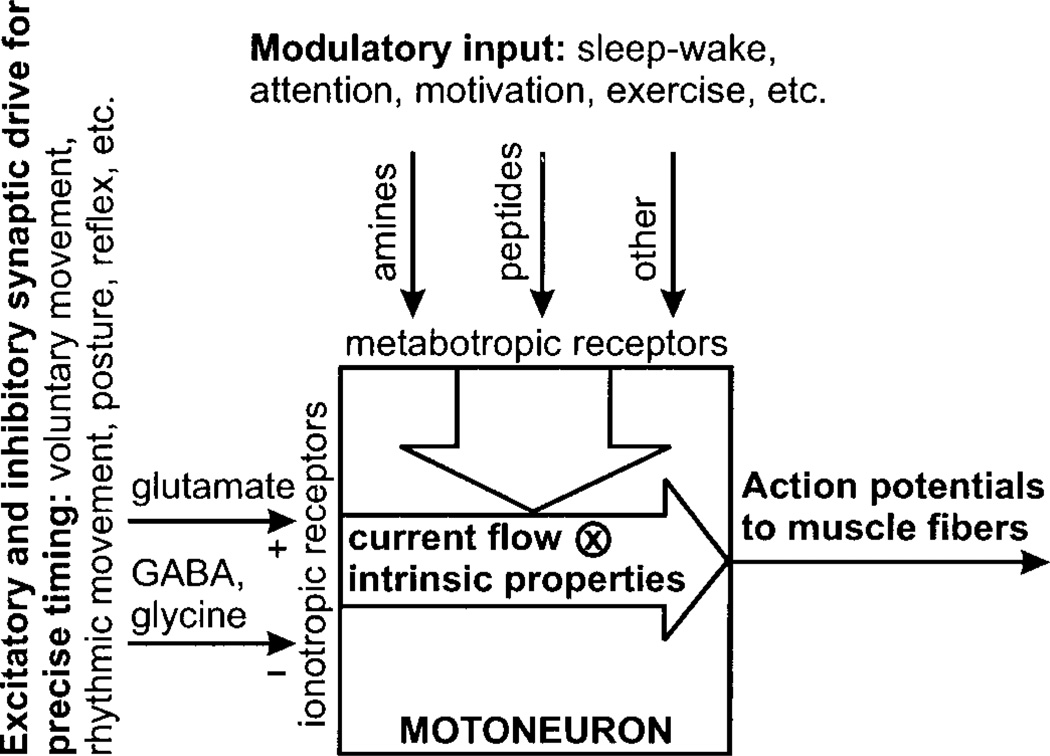
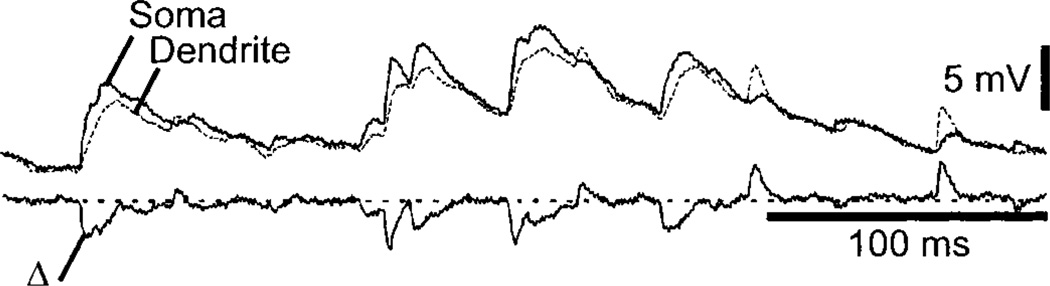
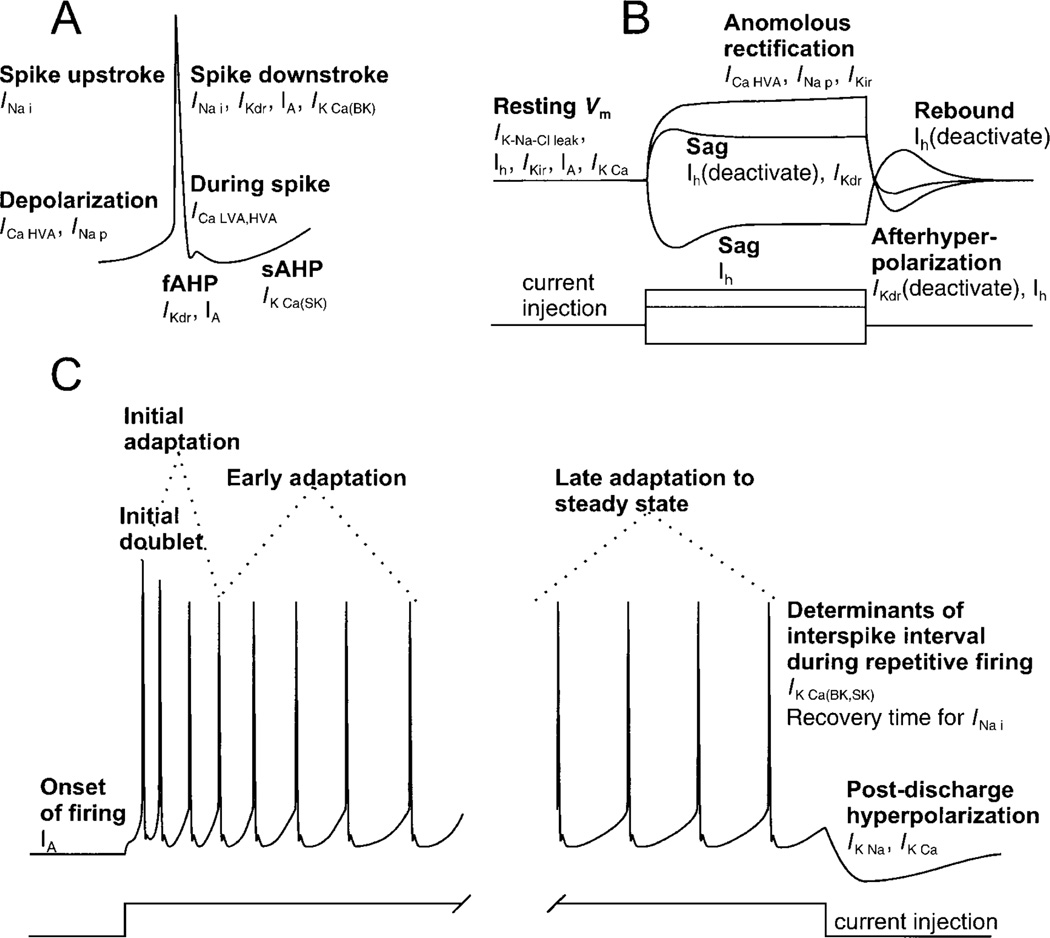
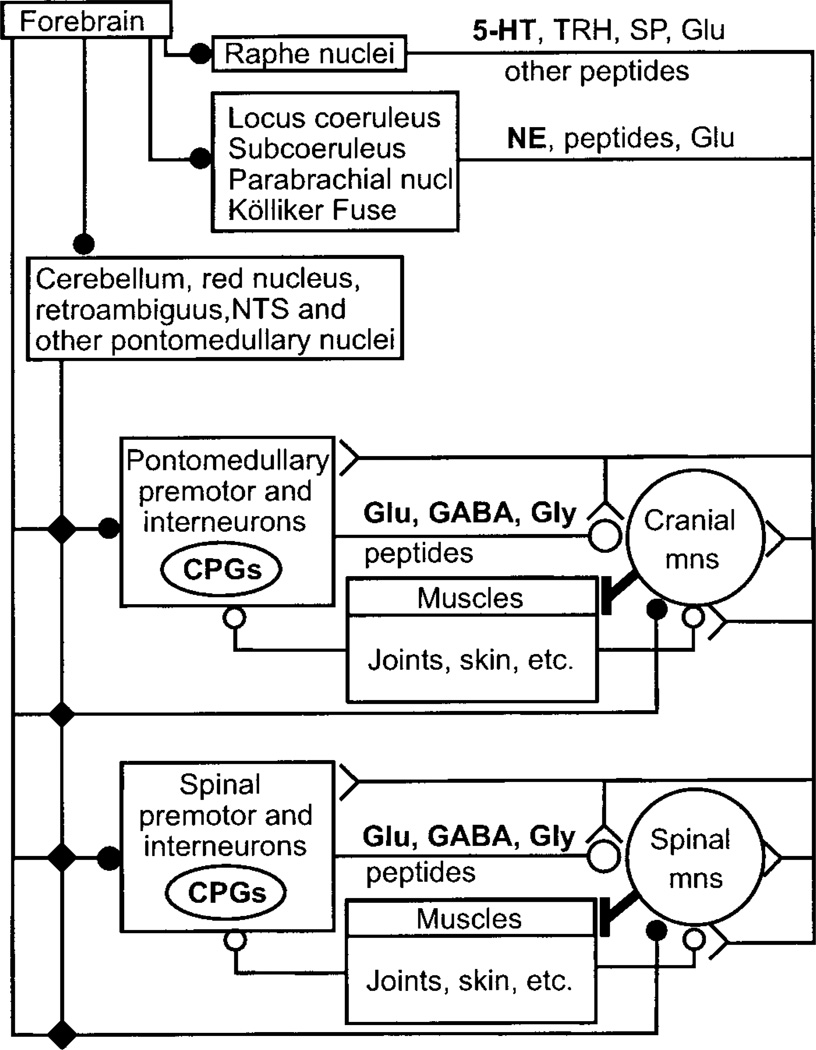
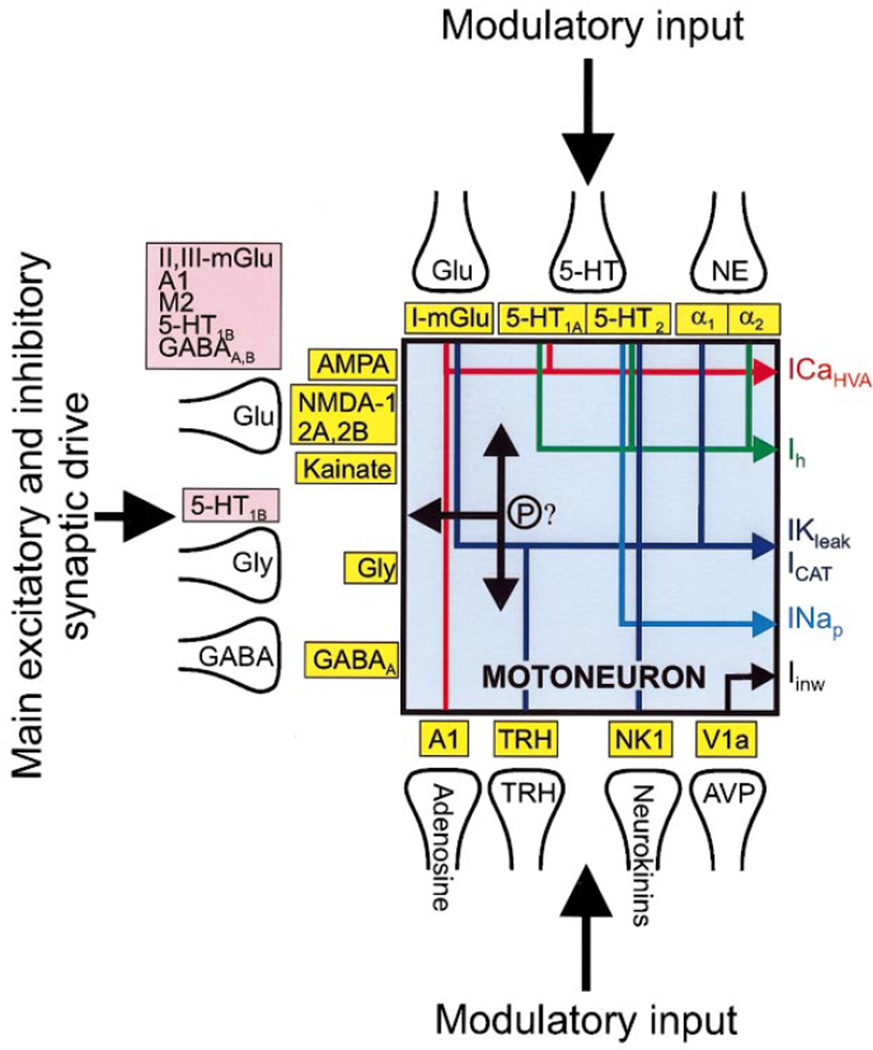
Movement, the fundamental component of behavior and the principal extrinsic action of the brain, is produced when skeletal muscles contract and relax in response to patterns of action potentials generated by motoneurons. The processes that determine the firing behavior of motoneurons are therefore important in understanding the transformation of neural activity to motor behavior. Here, we review recent studies on the control of motoneuronal excitability, focusing on synaptic and cellular properties. We first present a background description of motoneurons: their development, anatomical organization, and membrane properties, both passive and active. We then describe the general anatomical organization of synaptic input to motoneurons, followed by a description of the major transmitter systems that affect motoneuronal excitability, including ligands, receptor distribution, pre- and postsynaptic actions, signal transduction, and functional role. Glutamate is the main excitatory, and GABA and glycine are the main inhibitory transmitters acting through ionotropic receptors. These amino acids signal the principal motor commands from peripheral, spinal, and supraspinal structures. Amines, such as serotonin and norepinephrine, and neuropeptides, as well as the glutamate and GABA acting at metabotropic receptors, modulate motoneuronal excitability through pre- and postsynaptic actions. Acting principally via second messenger systems, their actions converge on common effectors, e.g., leak K(+) current, cationic inward current, hyperpolarization-activated inward current, Ca(2+) channels, or presynaptic release processes. Together, these numerous inputs mediate and modify incoming motor commands, ultimately generating the coordinated firing patterns that underlie muscle contractions during motor behavior.
Figures
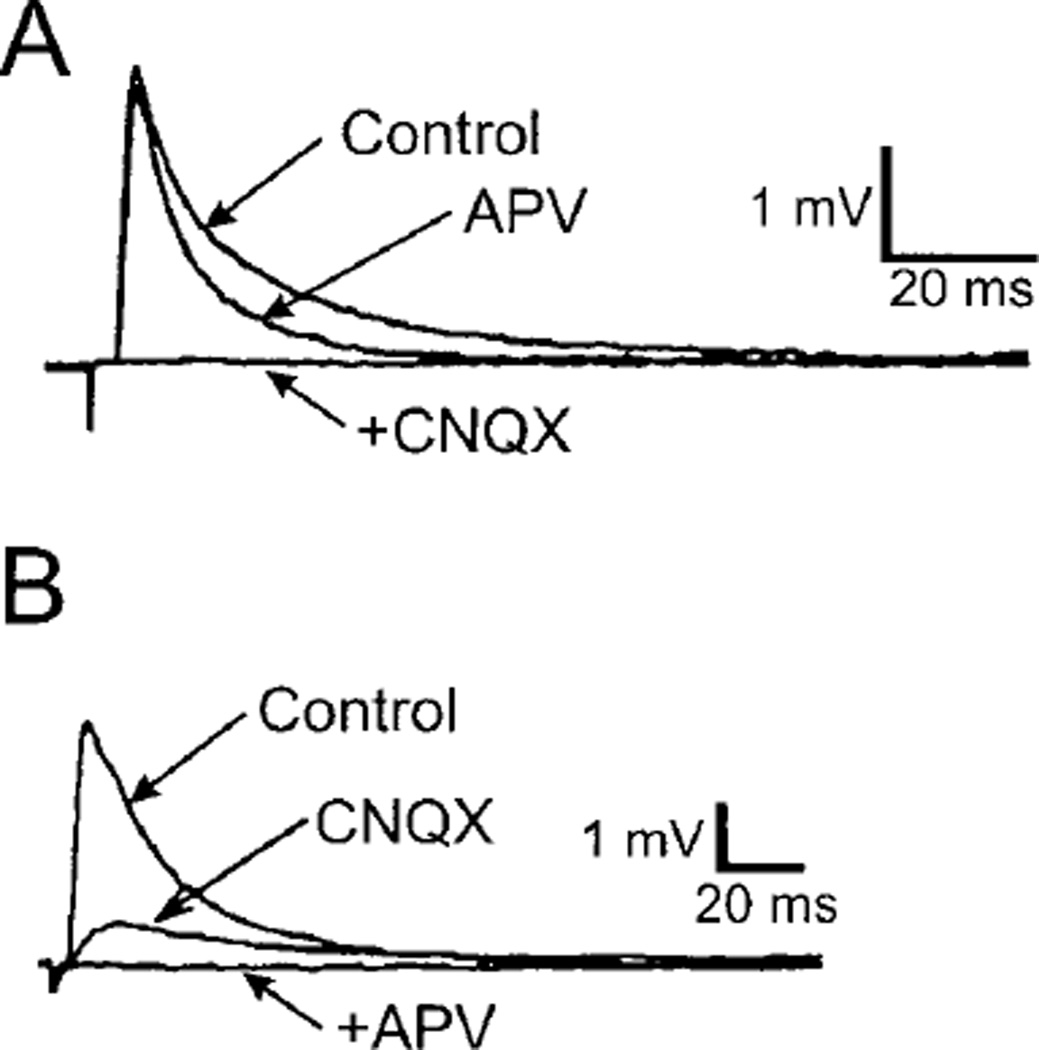
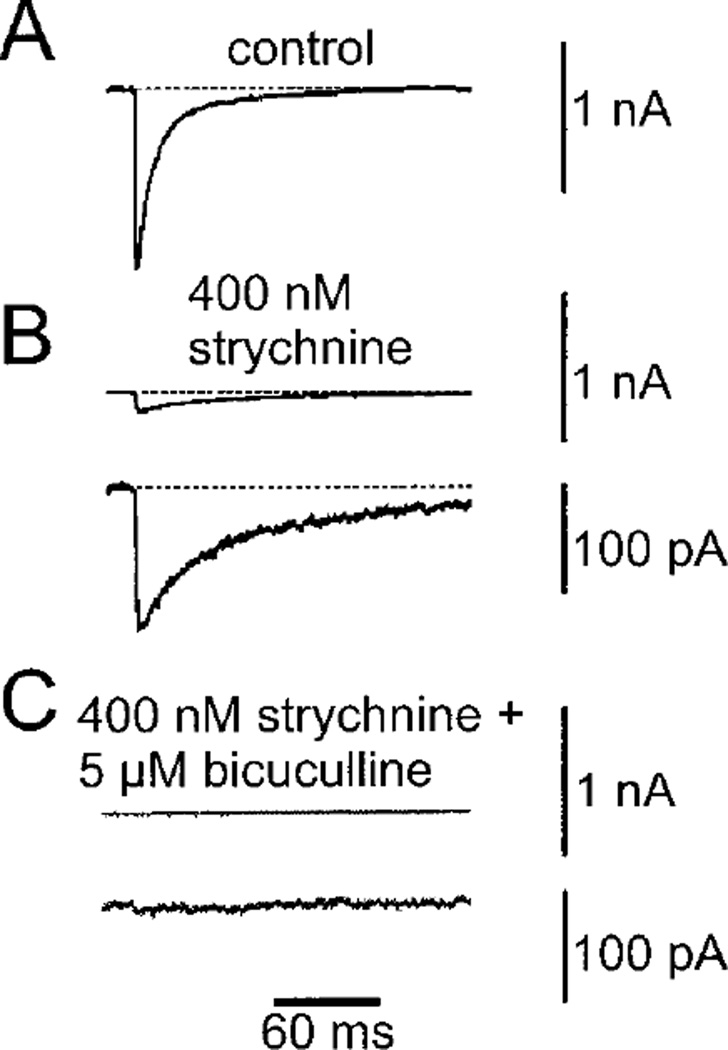
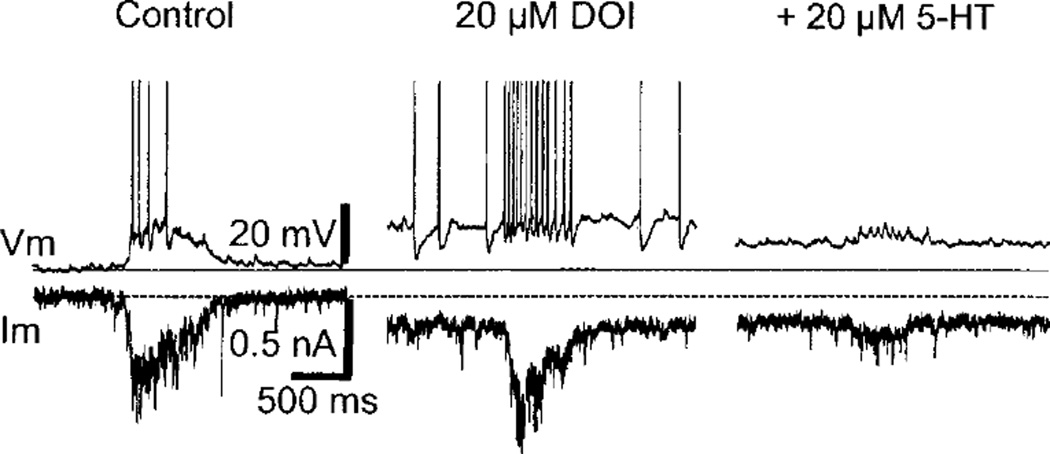
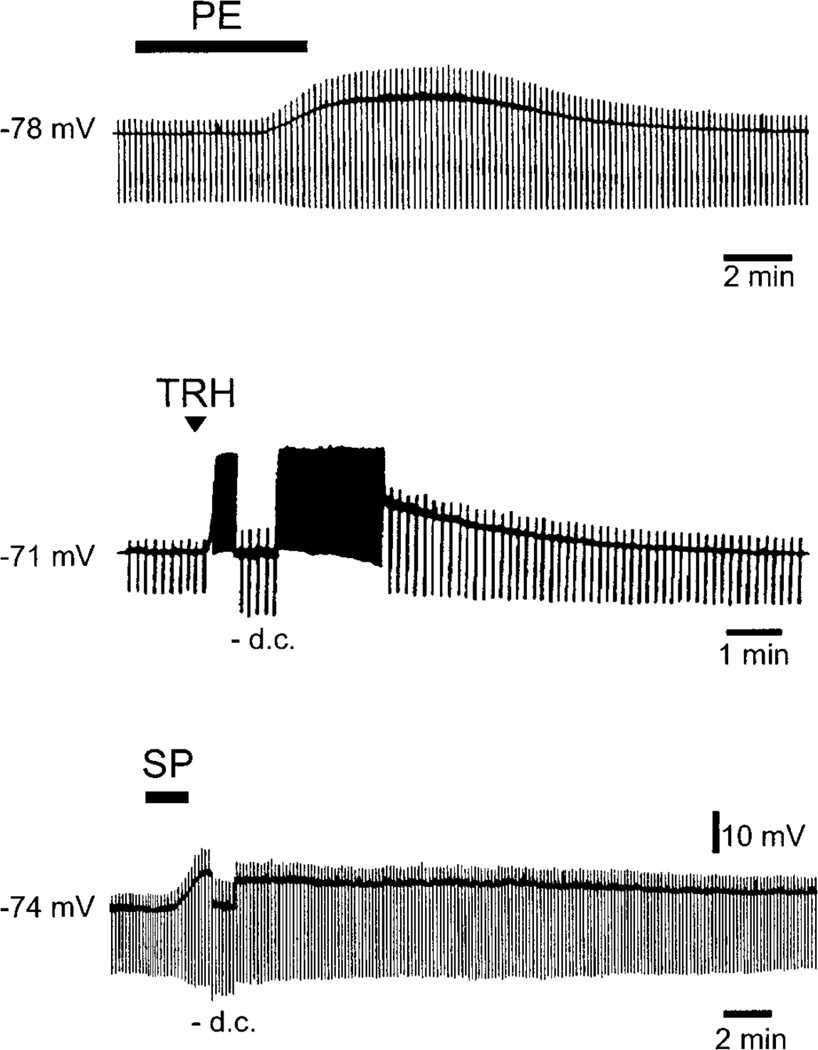
References
-
- Abbracchio MP, Burnstock G. Purinoceptors: are there families of P2X and P2Y purinoceptors? Pharmacol. Ther. 1994;64:445–475. - PubMed
-
- Abbracchio MP, Burnstock G. Purinergic signalling: pathophysiological roles. Jpn. J. Physiol. 1998;78:113–145. - PubMed
-
- Abe T, Sugihara H, Nawa H, Shigemoto R, Mizuno N, Nakanishi S. Molecular characterization of a novel metabotropic glutamate receptor mGluR5 coupled to inositol phosphate/Ca2+ signal transduction. J Biol. Chem. 1992;267:13361–13368. - PubMed
-
- Aghajanian GK. Modulation of a transient outward current in serotonergic neurones by alpha 1-adrenoceptors. Nature. 1985;315:501–503. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous