

Gene 33/Mig-6, a transcriptionally inducible adapter protein that binds GTP-Cdc42 and activates SAPK/JNK. A potential marker transcript for chronic pathologic conditions, such as diabetic nephropathy. Possible role in the response to persistent stress
- PMID: 10749885
- PMCID: PMC3668664
- DOI: 10.1074/jbc.M909735199
Gene 33/Mig-6, a transcriptionally inducible adapter protein that binds GTP-Cdc42 and activates SAPK/JNK. A potential marker transcript for chronic pathologic conditions, such as diabetic nephropathy. Possible role in the response to persistent stress
Abstract
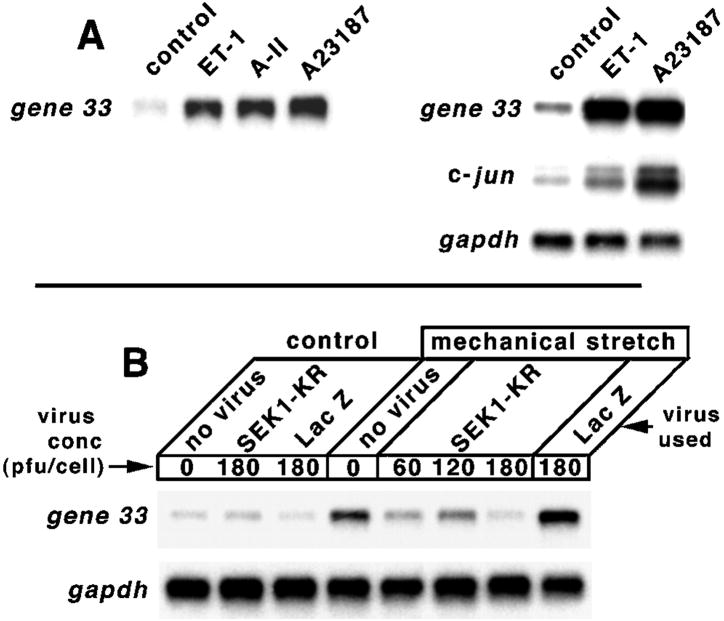
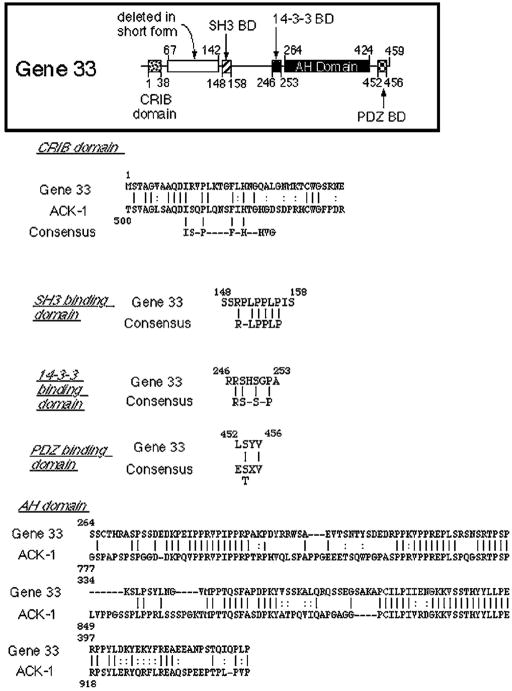
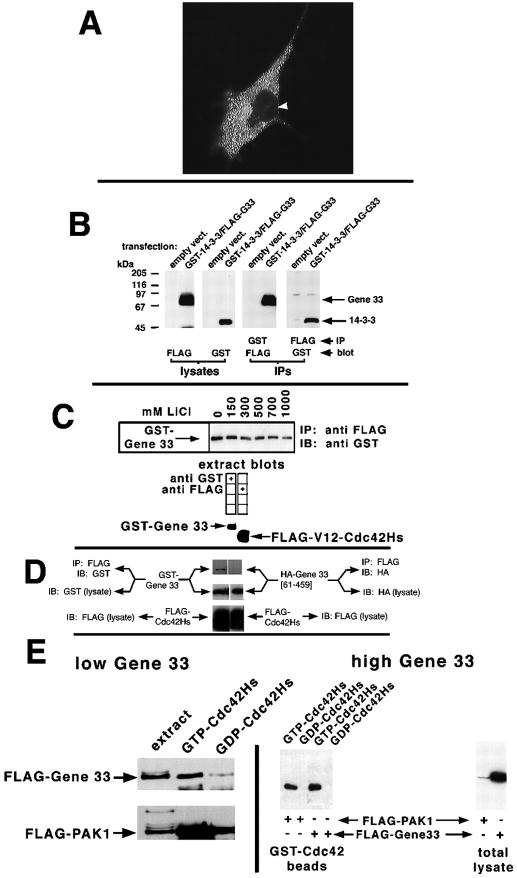
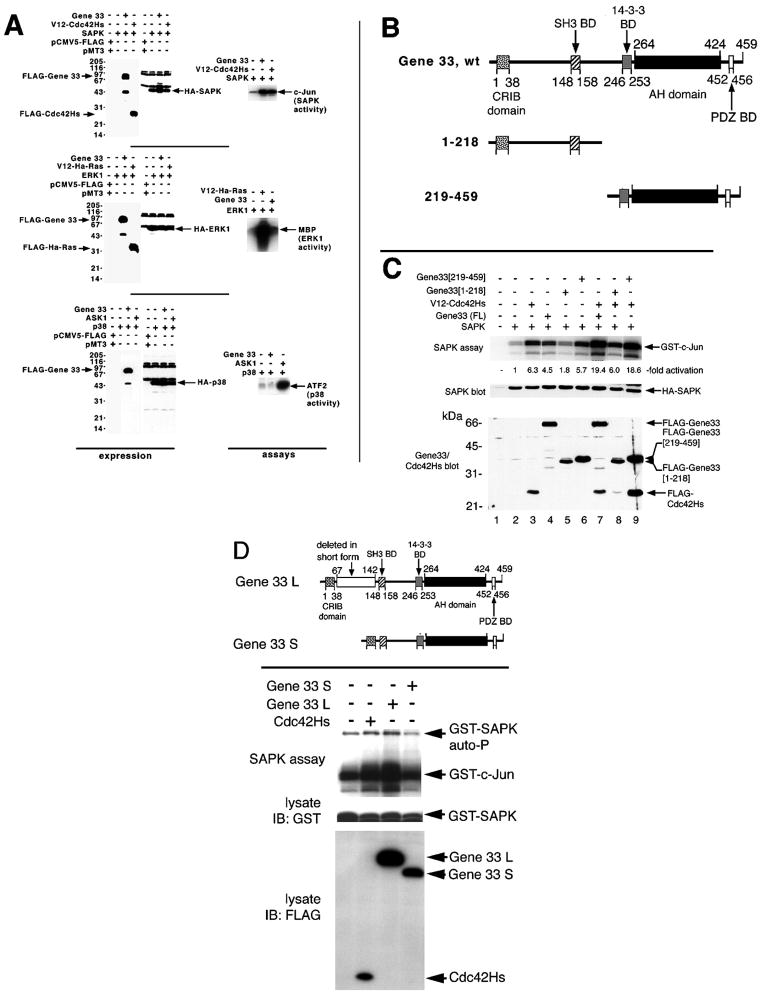
Chronic stresses, including the mechanical strain caused by hypertension or excess pulmonary ventilation pressure, lead to important clinical consequences, including hypertrophy and acute respiratory distress syndrome. Pathologic hypertrophy contributes to decreased organ function and, ultimately, organ failure; and cardiac and diabetic renal hypertrophy are major causes of morbidity and morality in the developed world. Likewise, acute respiratory distress syndrome is a serious potential side effect of mechanical pulmonary ventilation. Whereas the deleterious effects of chronic stress are well established, the molecular mechanisms by which these stresses affect cell function are still poorly characterized. gene 33 (also called mitogen-inducible gene-6, mig-6) is an immediate early gene that is transcriptionally induced by a divergent array of extracellular stimuli. The physiologic function of Gene 33 is unknown. Here we show that gene 33 mRNA levels increase sharply in response to a set of commonly occurring chronic stress stimuli: mechanical strain, vasoactive peptides, and diabetic nephropathy. Induction of gene 33 requires the stress-activated protein kinases (SAPKs)/c-Jun NH(2)-terminal kinases. This expression pattern suggests that gene 33 is a potential marker for diabetic nephropathy and other pathologic responses to persistent sublethal stress. The structure of Gene 33 indicates an adapter protein capable of binding monomeric GTPases of the Rho subfamily. Consistent with this, Gene 33 interacts in vivo and, in a GTP-dependent manner, in vitro with Cdc42Hs; and transient expression of Gene 33 results in the selective activation of the SAPKs. These results imply a reciprocal, positive feedback relationship between Gene 33 expression and SAPK activation. Expression of Gene 33 at sufficient levels may enable a compensatory reprogramming of cellular function in response to chronic stress, which may have pathophysiological consequences.
Figures



References
-
- Fine LG, Norman JT, Kujubu DA, Knecht A. In: The Kidney: Physiology and Pathophysiology. Seldin DW, Geibisch G, editors. Raven Press; New York: 1992. pp. 3113–3134.
-
- Hunter JJ, Chien KR. N Engl J Med. 1999;341:1276–1283. - PubMed
-
- Wyncoll DL, Evand TW. Lancet. 1999;354:497–501. - PubMed
-
- Kyriakis JM, Avruch J. J Biol Chem. 1996;271:24313–24316. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
