

A novel helicase-type protein in the nucleolus: protein NOH61
- PMID: 10749921
- PMCID: PMC14838
- DOI: 10.1091/mbc.11.4.1153
A novel helicase-type protein in the nucleolus: protein NOH61
Abstract
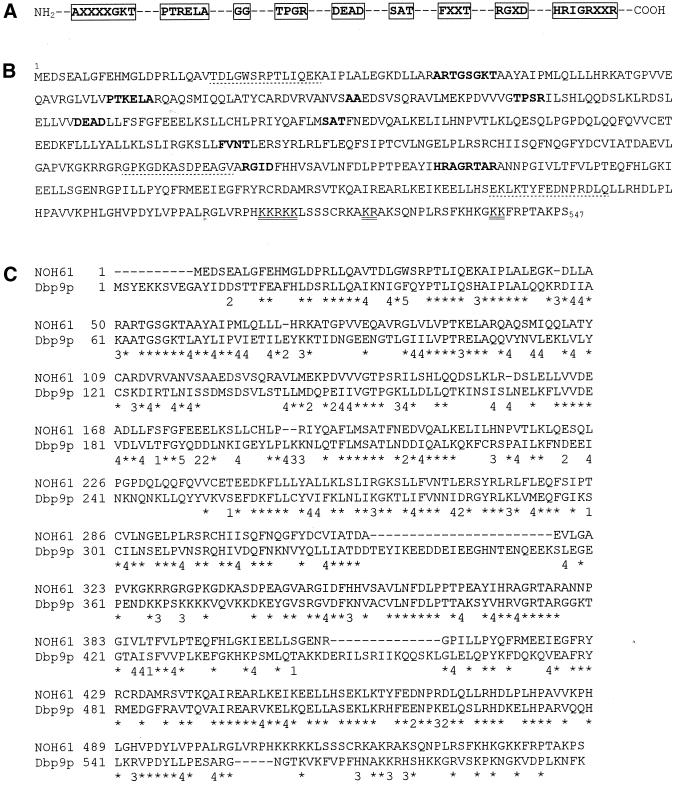
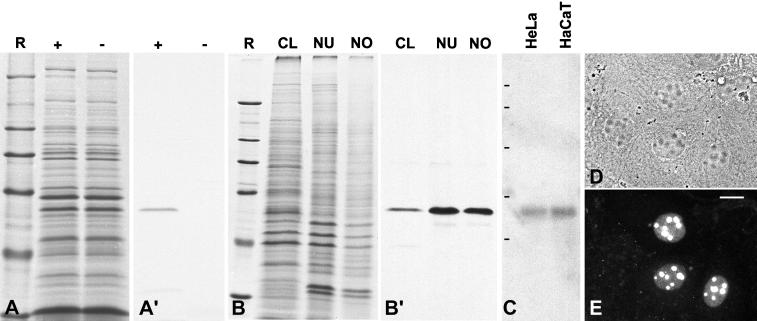
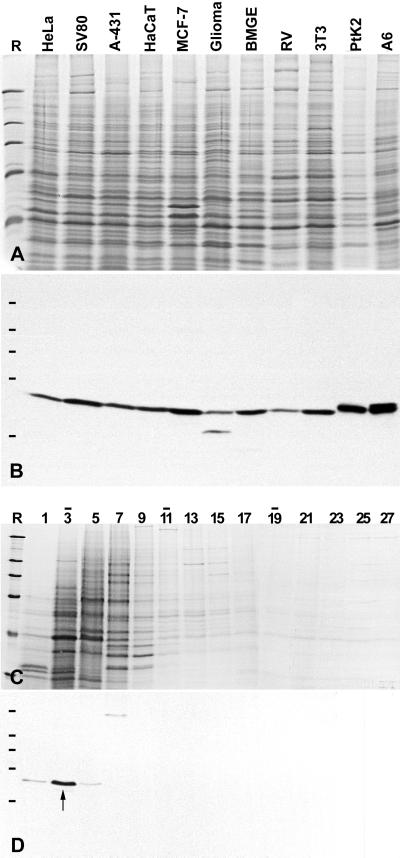
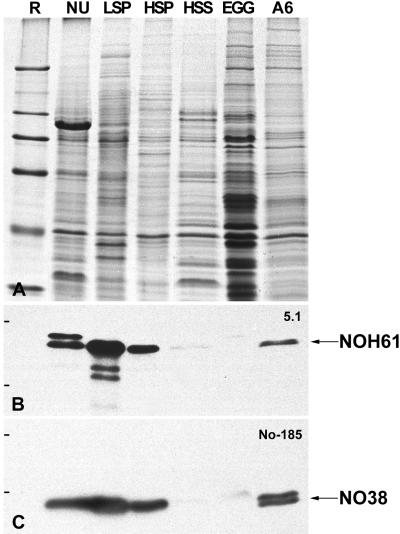
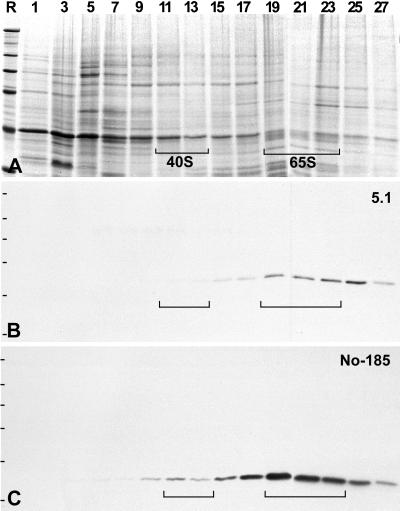
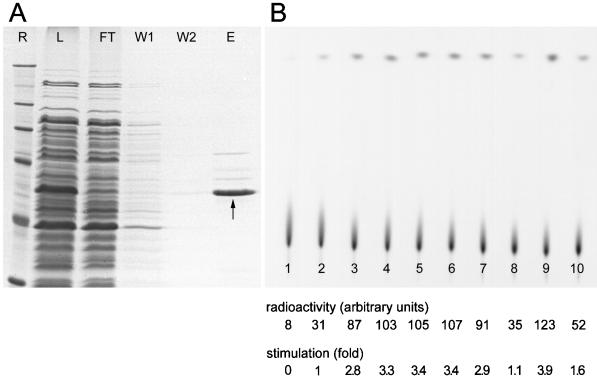
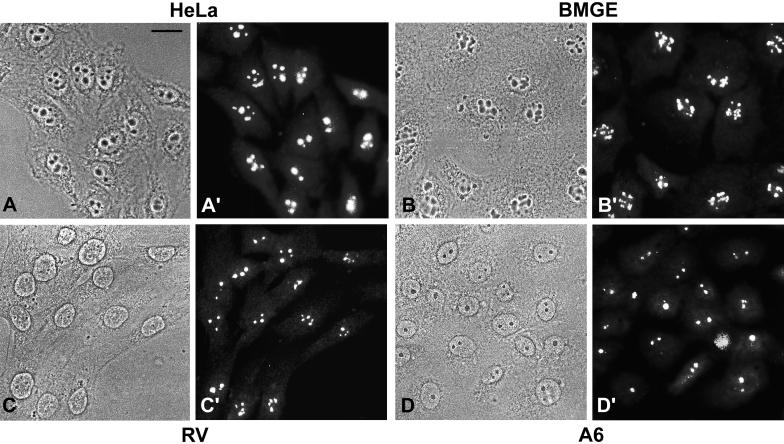
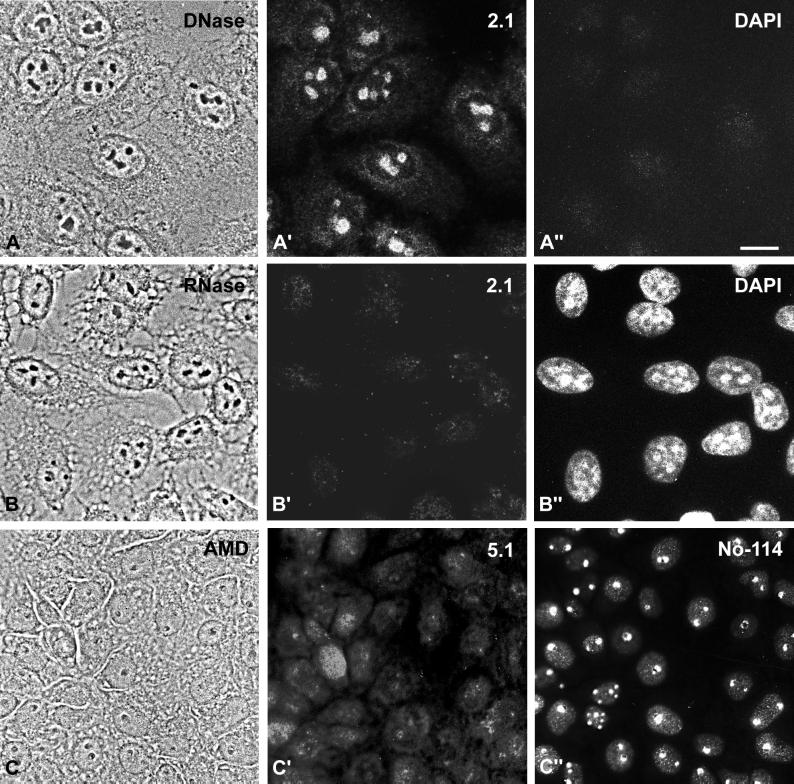
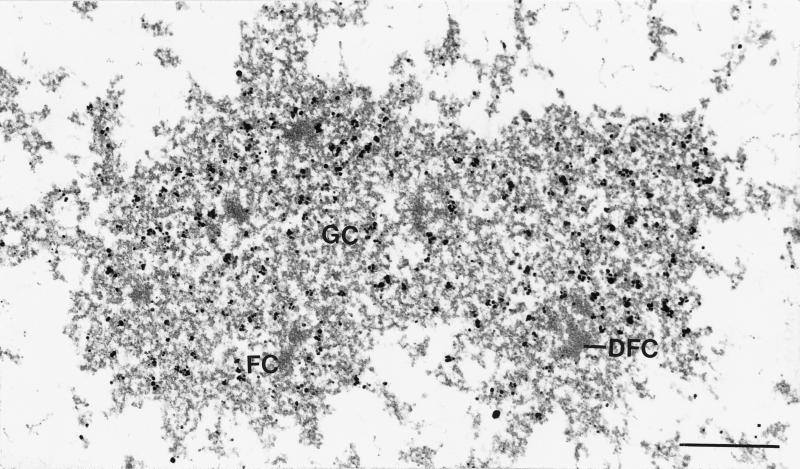
We report the identification, cDNA cloning, and molecular characterization of a novel, constitutive nucleolar protein. The cDNA-deduced amino acid sequence of the human protein defines a polypeptide of a calculated mass of 61.5 kDa and an isoelectric point of 9.9. Inspection of the primary sequence disclosed that the protein is a member of the family of "DEAD-box" proteins, representing a subgroup of putative ATP-dependent RNA helicases. ATPase activity of the recombinant protein is evident and stimulated by a variety of polynucleotides tested. Immunolocalization studies revealed that protein NOH61 (nucleolar helicase of 61 kDa) is highly conserved during evolution and shows a strong accumulation in nucleoli. Biochemical experiments have shown that protein NOH61 synthesized in vitro sediments with approximately 11.5 S, i.e., apparently as homo-oligomeric structures. By contrast, sucrose gradient centrifugation analysis of cellular extracts obtained with buffers of elevated ionic strength (600 mM NaCl) revealed that the solubilized native protein sediments with approximately 4 S, suggestive of the monomeric form. Interestingly, protein NOH61 has also been identified as a specific constituent of free nucleoplasmic 65S preribosomal particles but is absent from cytoplasmic ribosomes. Treatment of cultured cells with 1) the transcription inhibitor actinomycin D and 2) RNase A results in a complete dissociation of NOH61 from nucleolar structures. The specific intracellular localization and its striking sequence homology to other known RNA helicases lead to the hypothesis that protein NOH61 might be involved in ribosome synthesis, most likely during the assembly process of the large (60S) ribosomal subunit.
Figures
References
-
- Alber T. Structure of the leucine zipper. Curr Opin Genet Dev. 1992;2:205–210. - PubMed
-
- Busch H. Nucleolar and nucleolonemal proteins of cancer cells. J Tumor Marker Oncol. 1997;12:4–68.
-
- Cairns C, McStay B. Identification and cDNA cloning of a Xenopus nucleolar phosphoprotein, xNopp180, that is the homolog of the rat nucleolar protein Nopp140. J Cell Sci. 1995;108:3339–3347. - PubMed
-
- Chan PK. Characterization and cellular localization of nucleophosmin/B23 in HeLa cells treated with selected cytotoxic agents (studies of B23-translocation mechanism) Exp Cell Res. 1992;203:174–181. - PubMed
-
- Chan PK, Chan FY. Nucleophosmin/B23 (NPM) oligomer is a major and stable entity in HeLa cells. Biochim Biophys Acta. 1995;1262:37–42. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
