

Differential fear conditioning induces reciprocal changes in the sensory responses of lateral amygdala neurons to the CS(+) and CS(-)
- PMID: 10753976
- PMCID: PMC311326
- DOI: 10.1101/lm.7.2.97
Differential fear conditioning induces reciprocal changes in the sensory responses of lateral amygdala neurons to the CS(+) and CS(-)
Abstract
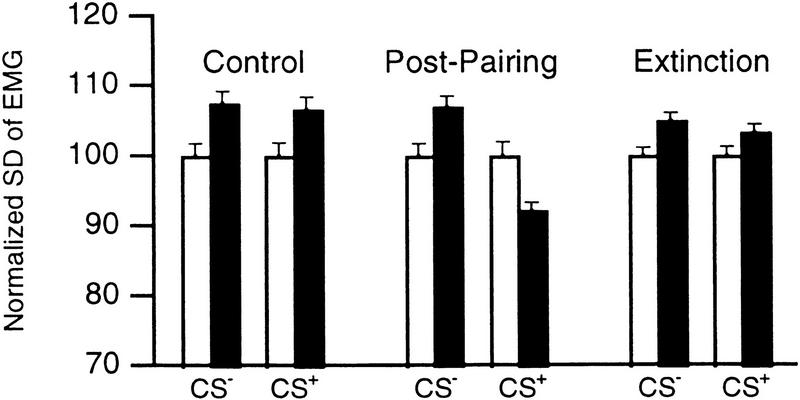
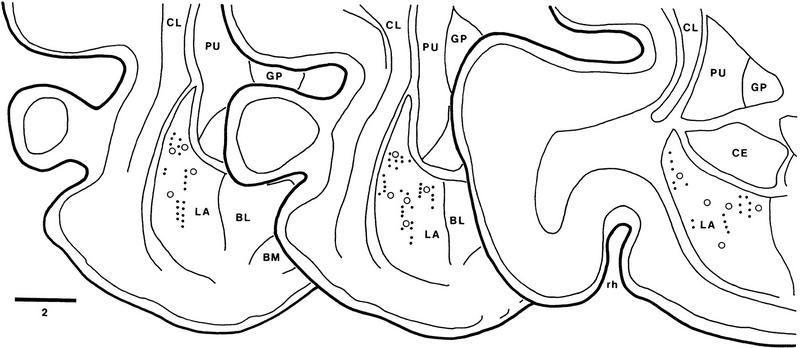
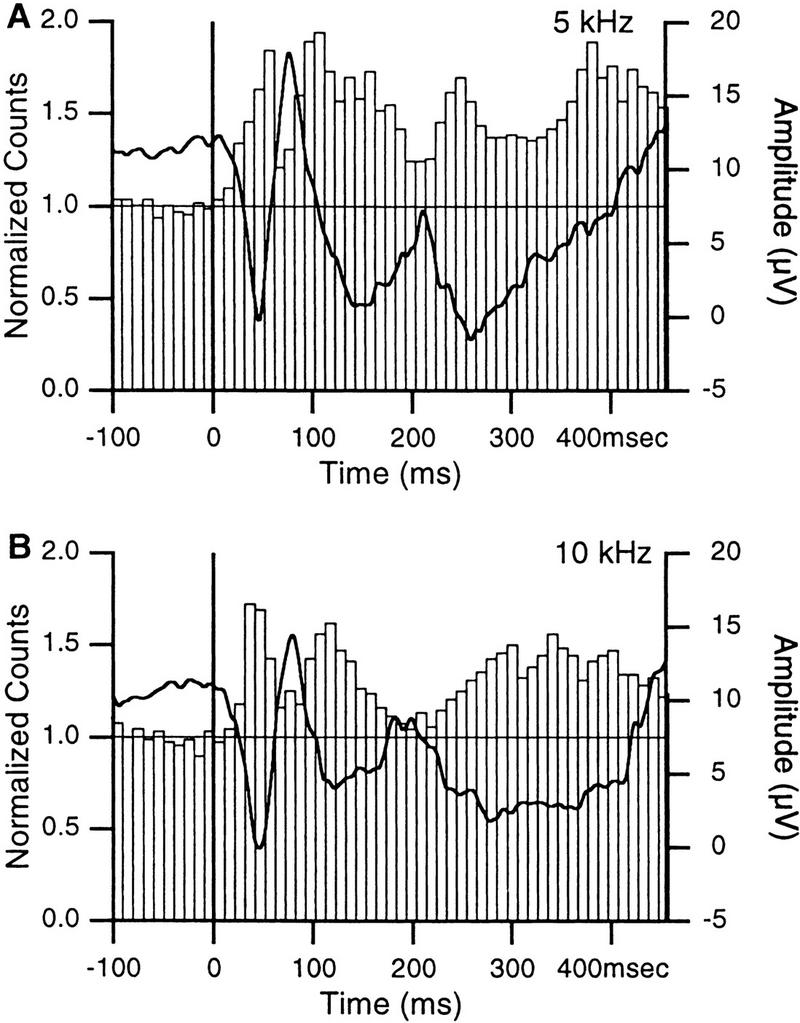
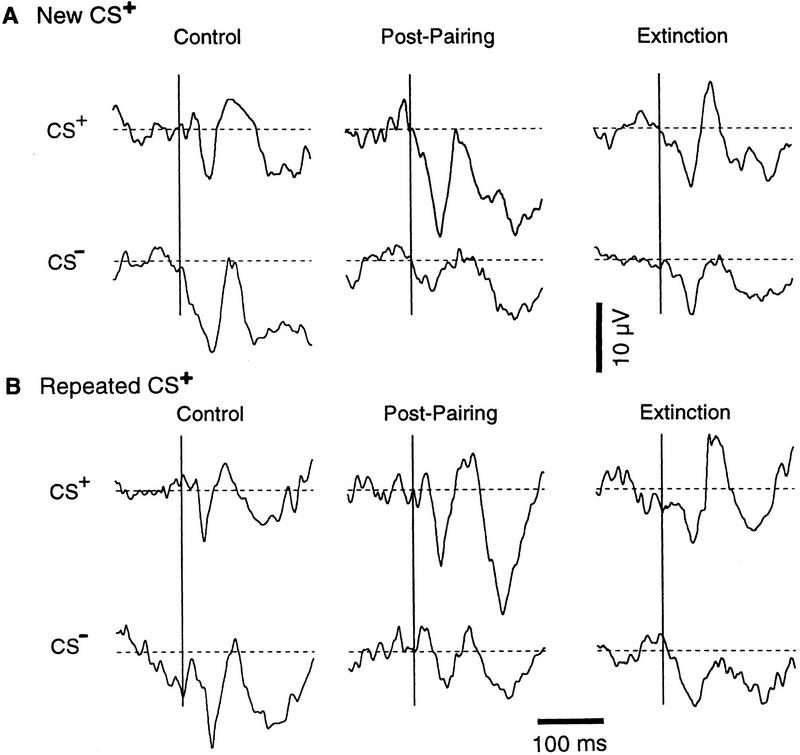
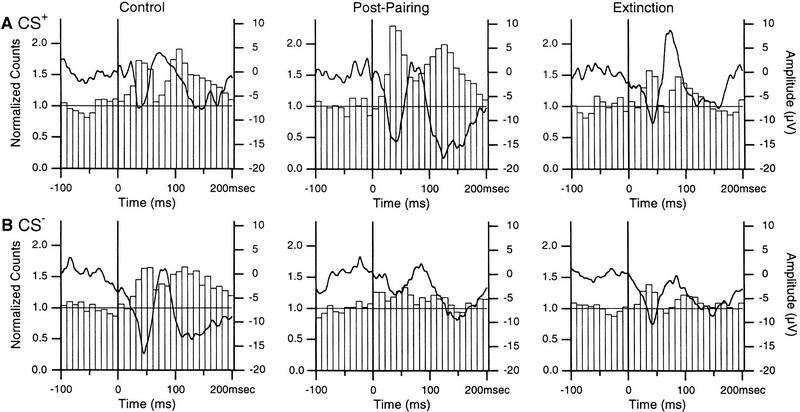
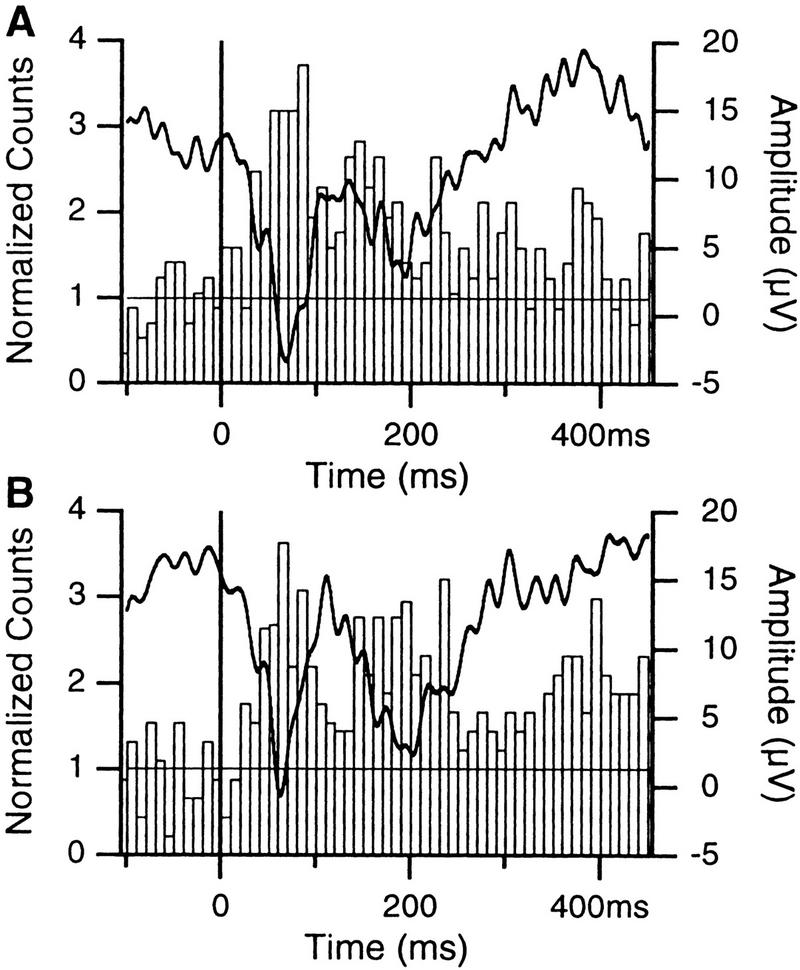
In classical fear conditioning, a neutral sensory stimulus (CS) acquires the ability to elicit fear responses after pairing to a noxious unconditioned stimulus (US). As amygdala lesions prevent the acquisition of fear responses and the lateral amygdaloid (LA) nucleus is the main input station of the amygdala for auditory afferents, the effect of auditory fear conditioning on the sensory responsiveness of LA neurons has been examined. Although conditioning was shown to increase CS-evoked LA responses, the specificity of the changes in responsiveness was not tested. Because conditioning might induce nonspecific increases in LA responses to auditory afferents, we re-examined this issue in conscious, head-restrained cats using a differential conditioning paradigm where only one of two tones (CS(+) but not CS(-)) was paired to the US. Differential conditioning increased unit and field responses to the CS(+), whereas responses to the CS(-) decreased. Such changes have never been observed in the amygdala except in cases where the CS(-) had been paired to the US before and fear responses not extinguished. This suggests that fear conditioning is not only accompanied by potentiation of amygdalopetal pathways conveying the CS(+) but also by the depression of sensory inputs unpaired to noxious stimuli.
Figures
References
-
- Amaral DG, Price JL, Pitkänen A, Carmichael ST. Anatomical organization of the primate amygdaloid complex. In: Aggleton JP, editor. The amygdala: Neurobiological aspects of emotion, memory, and mental dysfunction. New York, NY: Wiley-Liss; 1992. pp. 1–66.
-
- Bakin JS, Weinberger NM. Classical conditioning induces CS-specific receptive field plasticity in the auditory cortex of the guinea pig. Brain Res. 1990;536:271–286. - PubMed
-
- Bandler R. Induction of “rage” following microinjections of glutamate into midbrain but not hypothalamus of cats. Neurosci Lett. 1982;5:183–188. - PubMed
-
- Cahill L, Weinberger NM, Roozendaal B, McGaugh JL. Is the amygdala a locus of “conditioned fear”? Some questions and caveats. Neuron. 1999;23:227–228. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous