

The operant and the classical in conditioned orientation of Drosophila melanogaster at the flight simulator
- PMID: 10753977
- PMCID: PMC311324
- DOI: 10.1101/lm.7.2.104
The operant and the classical in conditioned orientation of Drosophila melanogaster at the flight simulator
Abstract
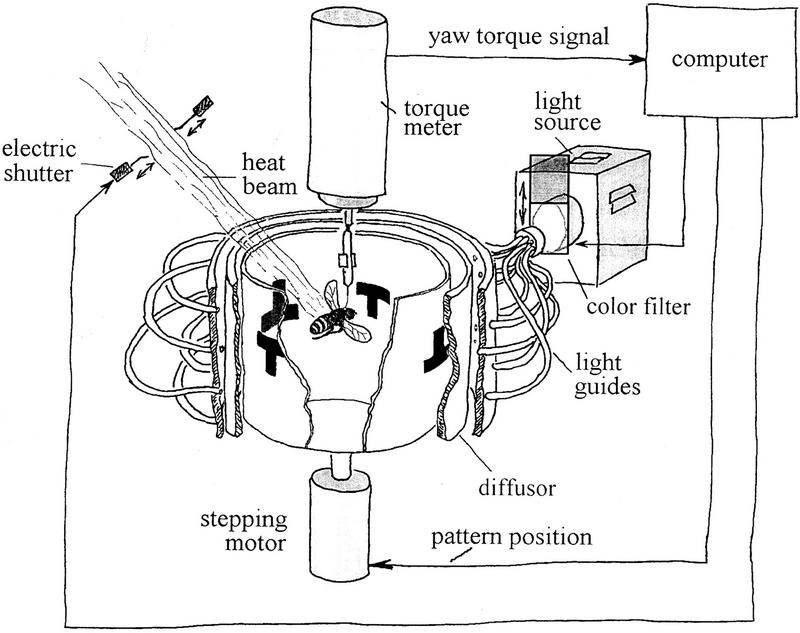
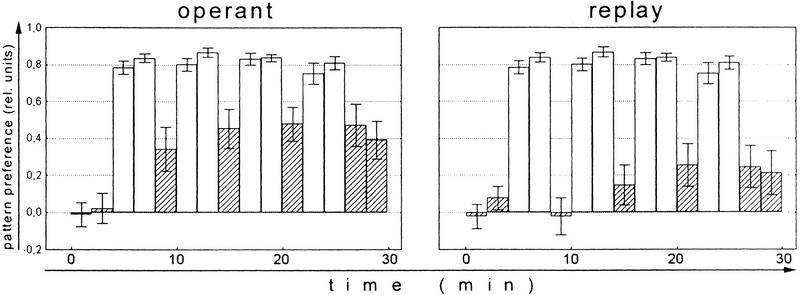
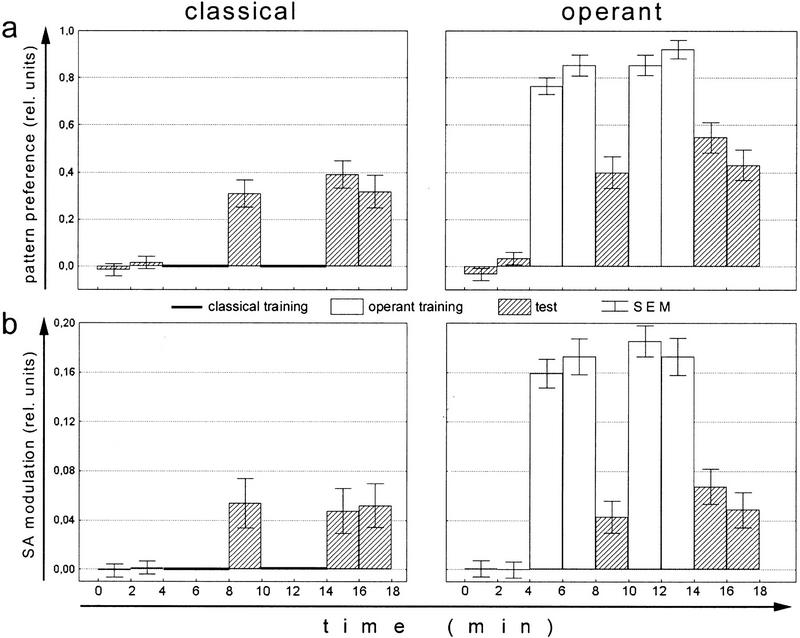
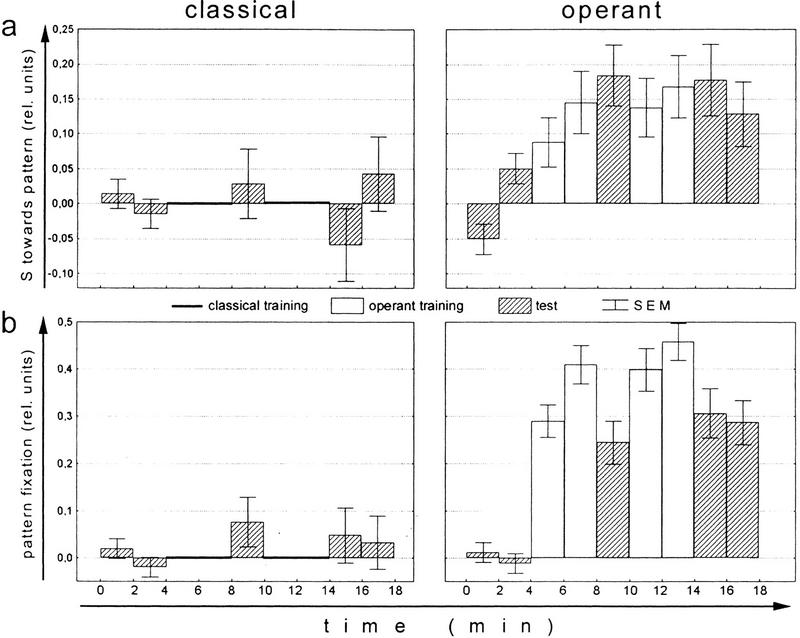
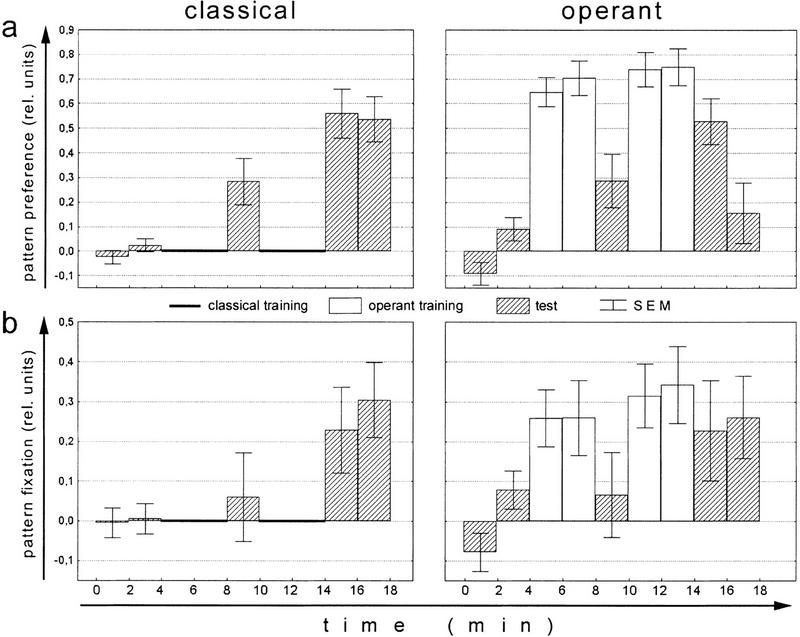
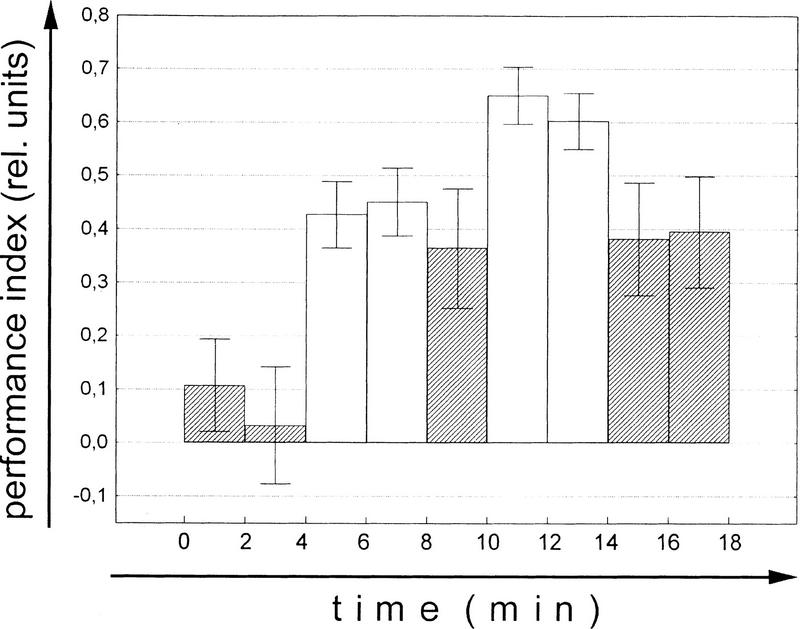
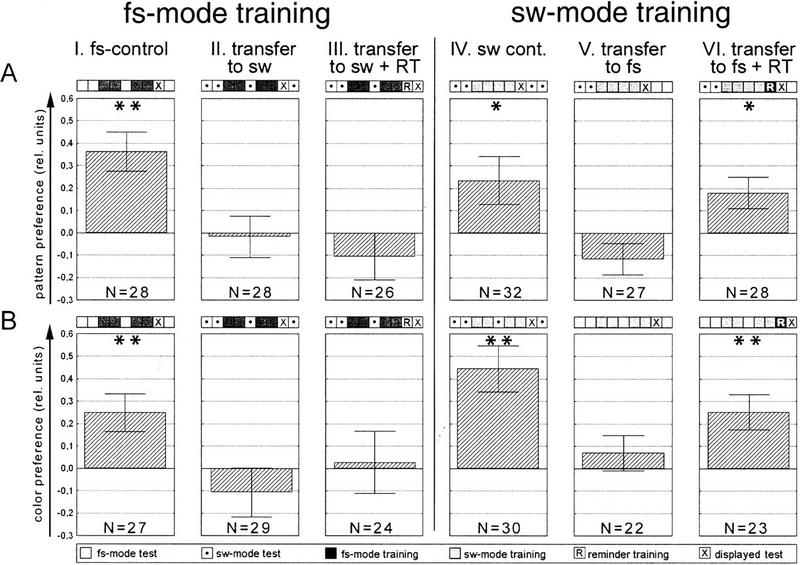
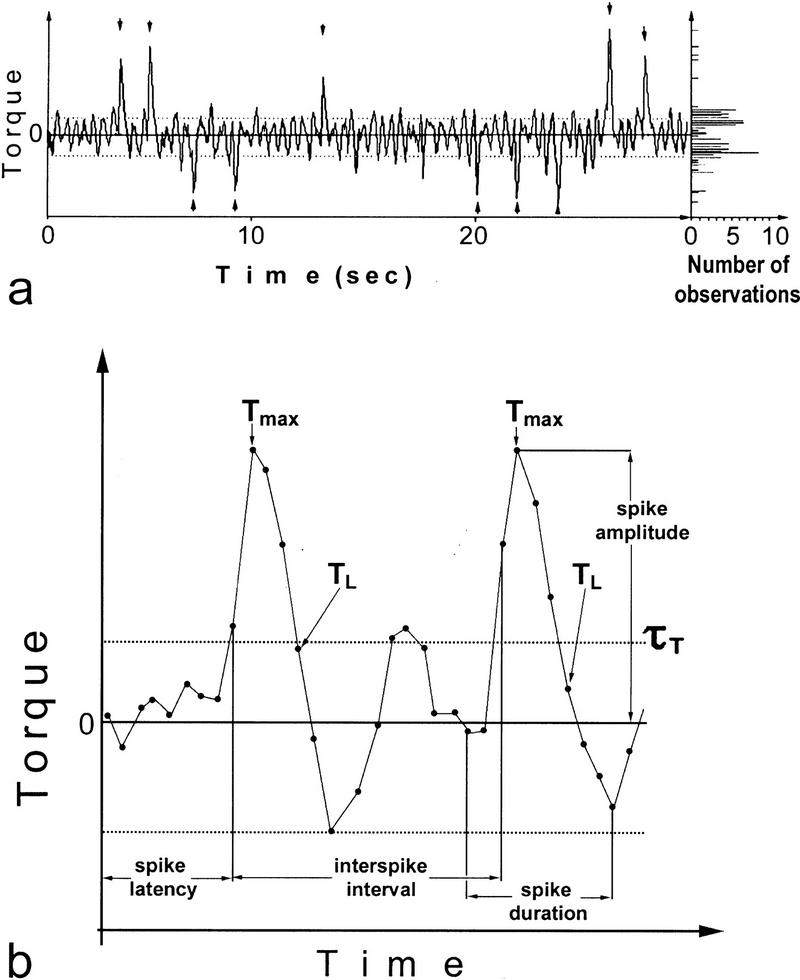
Ever since learning and memory have been studied experimentally, the relationship between operant and classical conditioning has been controversial. Operant conditioning is any form of conditioning that essentially depends on the animal's behavior. It relies on operant behavior. A motor output is called operant if it controls a sensory variable. The Drosophila flight simulator, in which the relevant behavior is a single motor variable (yaw torque), fully separates the operant and classical components of a complex conditioning task. In this paradigm a tethered fly learns, operantly or classically, to prefer and avoid certain flight orientations in relation to the surrounding panorama. Yaw torque is recorded and, in the operant mode, controls the panorama. Using a yoked control, we show that classical pattern learning necessitates more extensive training than operant pattern learning. We compare in detail the microstructure of yaw torque after classical and operant training but find no evidence for acquired behavioral traits after operant conditioning that might explain this difference. We therefore conclude that the operant behavior has a facilitating effect on the classical training. In addition, we show that an operantly learned stimulus is successfully transferred from the behavior of the training to a different behavior. This result unequivocally demonstrates that during operant conditioning classical associations can be formed.
Figures
References
-
- Balleine B. Asymmetrical interactions between thirst and hunger in Pavlovian-instrumental transfer. Q J Exp Psychol B. 1994;47:211–231. - PubMed
-
- Brembs B. “Classical and operant conditioning in Drosophila at the flight simulator.” Diploma Thesis. Germany: Julius-Maximilians-Universität Würzburg; 1996.
-
- Carew TJ, Sahley CL. Invertebrate learning and memory: From behavior to molecules. Annu Rev Neurosci. 1986;9:435–487. - PubMed
-
- Denniston JC, Miller RR, Matute H. Biological significance as determinant of cue competition. Psych Sci. 1996;7:325–331.
-
- Dill M, Wolf R, Heisenberg M. Visual pattern memory without shape recognition. Phil Trans R Soc Lond B. 1995;349:143–152. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases