

Review
doi: 10.1016/s0968-0004(00)01568-1.
Links between replication, recombination and genome instability in eukaryotes
Affiliations
- PMID: 10754554
- PMCID: PMC3635104
- DOI: 10.1016/s0968-0004(00)01568-1
Item in Clipboard
Review
Links between replication, recombination and genome instability in eukaryotes
Trends Biochem Sci.
2000 Apr.
Abstract
Double-strand breaks in DNA can be repaired by homologous recombination including break-induced replication. In this reaction, the end of a broken DNA invades an intact chromosome and primes DNA replication resulting in the synthesis of an intact chromosome. Break-induced replication has also been suggested to cause different types of genome rearrangements.
Figures

Two models for the repair of double-strand breaks (DSBs). (a) In this model, proposed by Szostak et al., the ends of the DSB (1) are digested by 5′→3′ nucleases (2). The resulting 3′ overhang invades another DNA molecule at a homologous region (3) generating a primer end that is extended by a DNA polymerase (4). The strand displaced by the propagation of the D-loop can pair with sequences from the other end of the break (3), forming a primer for repair synthesis (4), which is extended exclusively by leading-strand synthesis. Two Holliday junctions are formed (5) that can be resolved independently, yielding products without an associated crossover (6) of the flanking regions (if both Holliday junctions are resolved by cleaving in the same manner) or with (7) an associated crossover (when both Holliday junctions are resolved in an opposite manner). (b) In the break-induced replication (BIR) model, the primer generated by the invading 3′ end (3) directs leading-strand synthesis (4). Lagging-strand synthesis is initiated on the leading strand producing a double-stranded DNA molecule with identical information to that of the template chromosome (5,6).
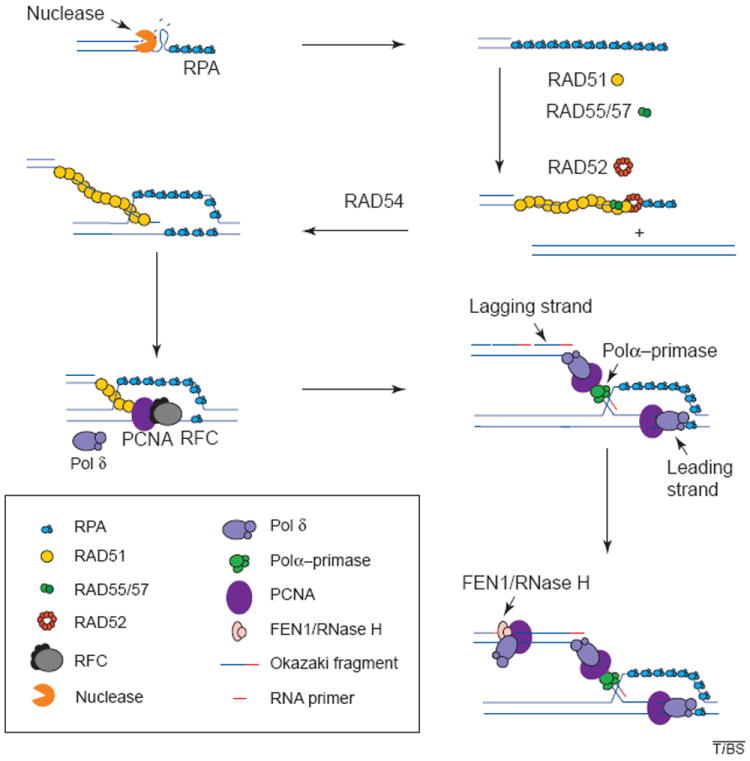
Recombination and associated DNA synthesis. The resection of the ends of the double strand creates a 3′ single-stranded DNA molecule. The binding of RPA to the single strand protects the DNA from degradation and disrupts any secondary structure. RAD51 is then loaded onto the DNA strand with the aid of RAD52, RAD55 and RAD57, and in the presence of RAD54, the RAD51 filament carries out strand exchange. The primer generated is recognized by the polymerase (Pol) δ accessory factor RFC, which loads PCNA onto the primer end generating a complex that is recognized by Pol δ or ε, which will carry out synthesis of the leading strand. Lagging-strand synthesis initiates when RNA primers are synthesized on the leading strand by the primase activity of the Pol-α–primase complex and are immediately extended by the Pol α into short DNAs. The Pol δ (or ε) system is then assembled on these DNA primers, replacing Pol α. The primers are then extended to yield Okazaki fragments. These accumulate on the lagging strand until they mature by the combined action of FEN1 and RNase H nucleases, which remove the RNA moiety of the fragment, leaving a gap that is then filled in by DNA Pol. DNA ligase seals the resulting nick yielding an intact double-stranded DNA molecule.

Mutagenic repair of double-strand breaks (DSBs). Broken chromosomes are repaired by break-induced replication. If tandem repeats (indicated as arrows) are present at the breakpoint and repair is initiated out of register, it can lead to deletions (1) of the sequences between the repeats. When the distal part of the chromosome is lost, the centromerecontaining fragment (with the centromere depicted as a solid black circle) can be repaired using another chromosome as the template leading to non-reciprocal translocations (2) where the broken chromosome acquires sequences from another chromosome (shown as gray shaded boxes) and in some cases it can be repaired using intrachromosomal regions (3). In all cases, the broken chromosome can capture telomere sequences (depicted as colored regions at the end of each chromosome) by BIR copying to the end of the template.
References
-
- Lett JT. Damage to DNA and chromatin structure from ionizing radiations, and the radiation sensitivities of mammalian cells. Prog Nucleic Acid Res Mol Biol. 1990;39:305–352. - PubMed
-
- Seigneur M, et al. RuvAB acts at arrested replication forks. Cell. 1998;30:419–430. - PubMed
-
- Haber JE. Meiosis: avoiding inappropriate relationships. Curr Biol. 1998;19:832–835. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
