

Intrahepatic induction of alpha/beta interferon eliminates viral RNA-containing capsids in hepatitis B virus transgenic mice
- PMID: 10756029
- PMCID: PMC111931
- DOI: 10.1128/jvi.74.9.4165-4173.2000
Intrahepatic induction of alpha/beta interferon eliminates viral RNA-containing capsids in hepatitis B virus transgenic mice
Abstract
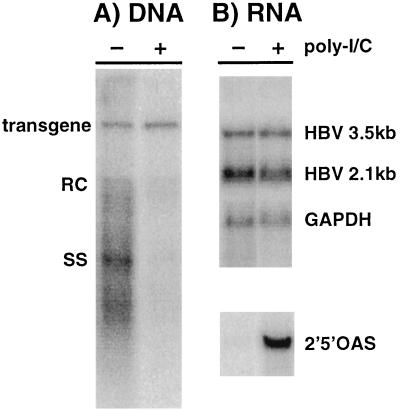
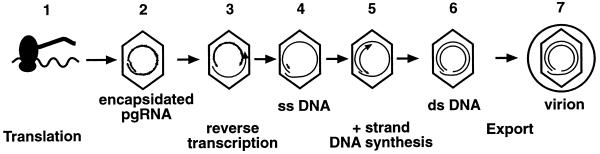
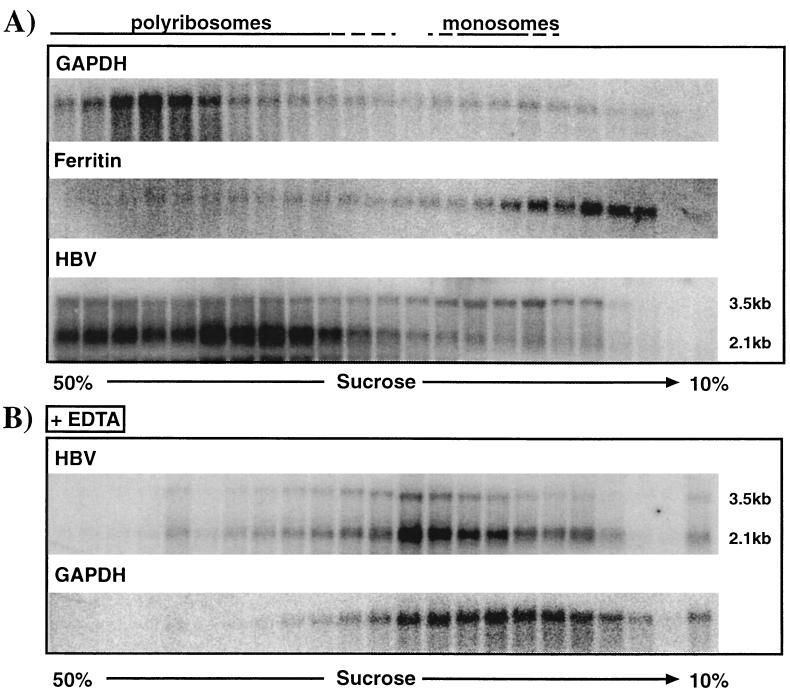
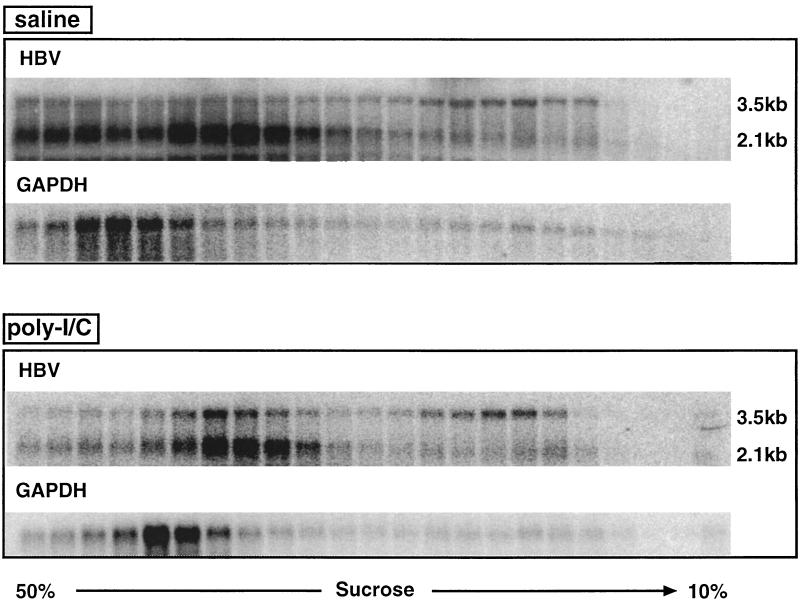
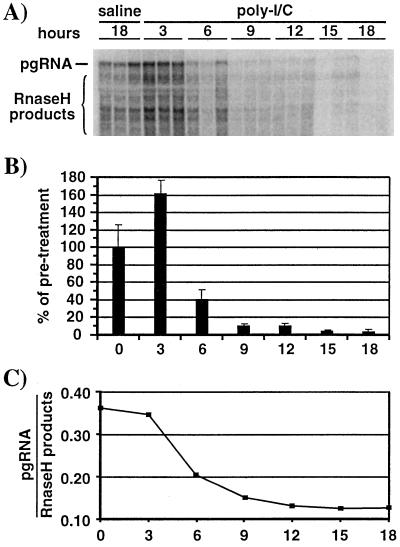
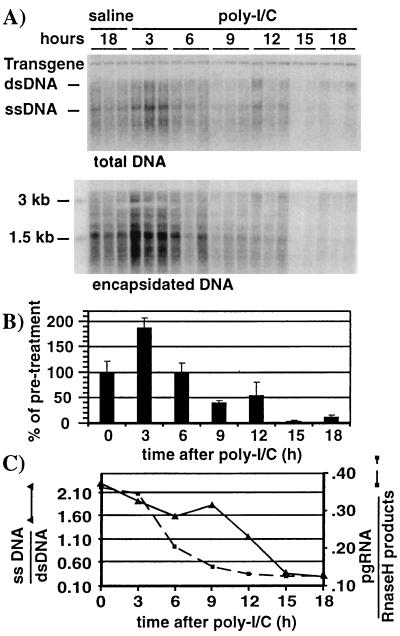
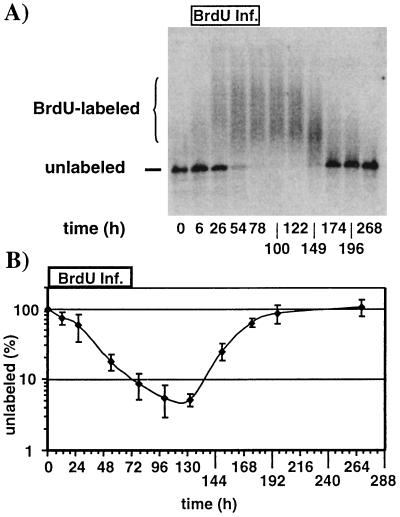
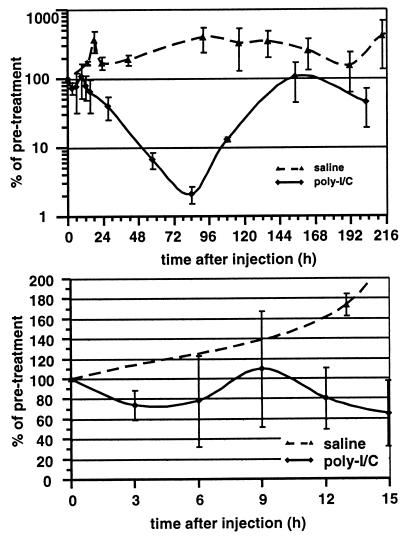
We have previously shown that hepatitis B virus (HBV) replication is abolished in the liver of HBV transgenic mice by stimuli that induce alpha/beta interferon (IFN-alpha/beta) in the liver. The present study was done to identify the step(s) in HBV replication that is affected by this cytokine in transgenic mice treated with the IFN-alpha/beta inducer polyinosinic-polycytidylic acid [poly(I-C)]. Here we show that the pool of cytoplasmic HBV pregenomic RNA (pgRNA)-containing capsids is reduced 10-fold within 9 h after poly(I-C) administration, while there is no change in the abundance of HBV mRNA or in the translational status of cytoplasmic HBV transcripts. In addition, we show that the pool of HBV DNA-containing capsids is not reduced to the same degree until at least 15 h posttreatment, and we show that virus export is not accelerated and the half-life of virions in the serum is unchanged. These results indicate that IFN-alpha/beta triggers intracellular events that either inhibit the assembly of pgRNA-containing capsids or accelerate their degradation, and that maturation and secretion of virus is responsible for clearance of HBV capsids and their cargo of replicative intermediates from the cytoplasm of the hepatocyte.
Figures
References
-
- Carroll S S, Chen E, Viscount T, Geib J, Sardana M K, Gehman J, Kuo L C. Cleavage of oligoribonucleotides by the 2′,5′-oligoadenylate-dependent ribonuclease L. J Biol Chem. 1996;271:4988–4992. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
