

Infectious bronchitis virus E protein is targeted to the Golgi complex and directs release of virus-like particles
- PMID: 10756047
- PMCID: PMC111949
- DOI: 10.1128/jvi.74.9.4319-4326.2000
Infectious bronchitis virus E protein is targeted to the Golgi complex and directs release of virus-like particles
Abstract
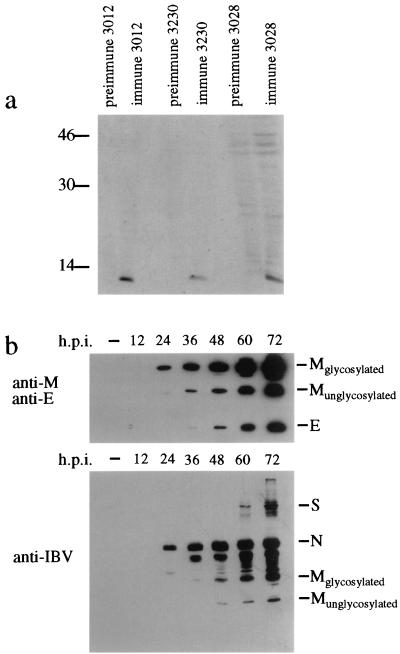
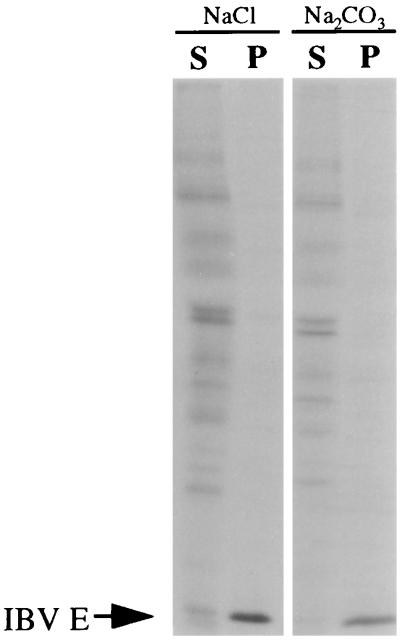
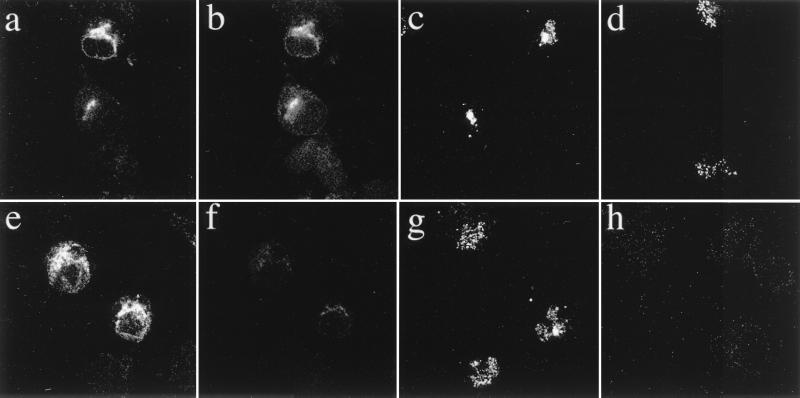
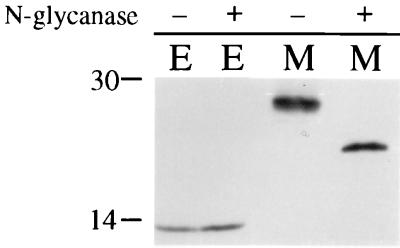
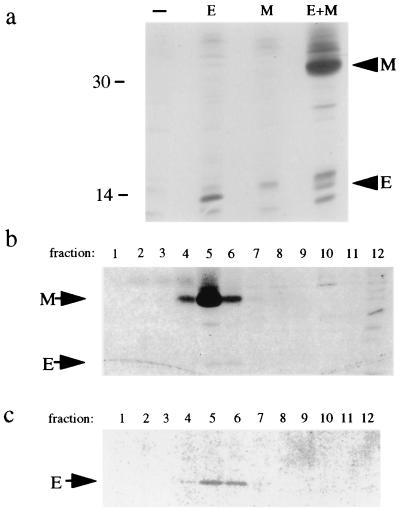
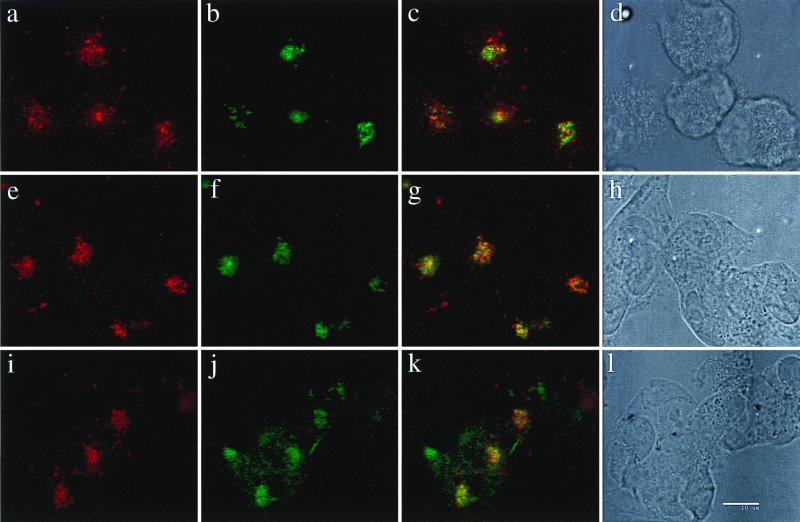
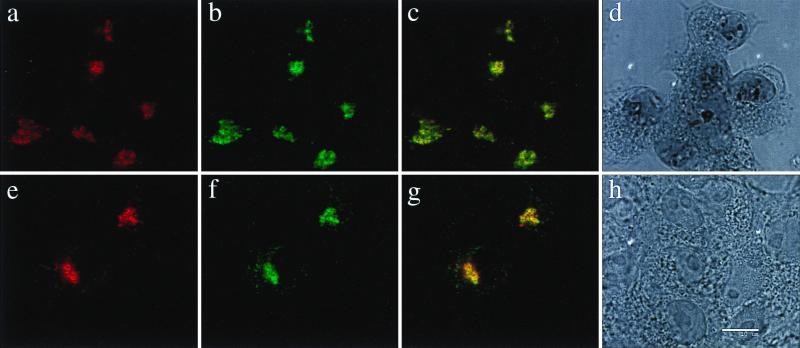
The coronavirus E protein is a poorly characterized small envelope protein present in low levels in virions. We are interested in the role of E in the intracellular targeting of infectious bronchitis virus (IBV) membrane proteins. We generated a cDNA clone of IBV E and antibodies to the E protein to study its cell biological properties in the absence of virus infection. We show that IBV E is an integral membrane protein when expressed in cells from cDNA. Epitope-specific antibodies revealed that the C terminus of IBV E is cytoplasmic and the N terminus is translocated. The short luminal N terminus of IBV E contains a consensus site for N-linked glycosylation, but the site is not used. When expressed using recombinant vaccinia virus, the IBV E protein is released from cells at low levels in sedimentable particles that have a density similar to that of coronavirus virions. The IBV M protein is incorporated into these particles when present. Indirect immunofluorescence microscopy showed that E is localized to the Golgi complex in cells transiently expressing IBV E. When coexpressed with IBV M, both from cDNA and in IBV infection, the two proteins are colocalized in Golgi membranes, near the coronavirus budding site. Thus, even though IBV E is present at low levels in virions, it is apparently expressed at high levels in infected cells near the site of virus assembly.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
